

Abstract
Background: Previous research in patients with anorexia nervosa showed heightened brain response during a taste reward conditioning task and heightened sensitivity to rewarding and punishing stimuli. Here we tested the hypothesis that individuals recovered from anorexia nervosa would also experience greater brain activation during this task as well as higher sensitivity to salient stimuli than controls.
Methods: Women recovered from restricting-type anorexia nervosa and healthy control women underwent fMRI during application of a prediction error taste reward learning paradigm.
Results: Twenty-four women recovered from anorexia nervosa (mean age 30.3 ± 8.1 yr) and 24 control women (mean age 27.4 ± 6.3 yr) took part in this study. The recovered anorexia nervosa group showed greater left posterior insula activation for the prediction error model analysis than the control group (family-wise error– and small volume–corrected p < 0.05). A group × condition analysis found greater posterior insula response in women recovered from anorexia nervosa than controls for unexpected stimulus omission, but not for unexpected receipt. Sensitivity to punishment was elevated in women recovered from anorexia nervosa.
Limitations: This was a cross-sectional study, and the sample size was modest.
Conclusion: Anorexia nervosa after recovery is associated with heightened prediction error–related brain response in the posterior insula as well as greater response to unexpected reward stimulus omission. This finding, together with behaviourally increased sensitivity to punishment, could indicate that individuals recovered from anorexia nervosa are particularly responsive to punishment. The posterior insula processes somatosensory stimuli, including unexpected bodily states, and greater response could indicate altered perception or integration of unexpected or maybe unwanted bodily feelings. Whether those findings develop during the ill state or whether they are biological traits requires further study.
Introduction
Anorexia nervosa is a psychiatric disorder characterized by intense fear of weight gain and persistent restriction of energy intake leading to significantly low body weight.1 While little continues to be understood regarding brain pathways that drive anorexia nervosa–related behaviours, an important theme to emerge is the neural representation of reward of food and nonfood stimuli.
The majority of functional brain imaging studies that tested brain response to visual food cues in individuals with eating disorders, including anorexia nervosa, have suggested altered limbic and reward processing regions as well as altered function in frontal cortical areas that contribute to cognitive control,2 while another review highlighted that those cues were typically associated with brain activations that indicated high anxiety.3 Some studies applied complex taste stimuli, such as chocolate milk, and those results of greater activation in the amygdala in individuals with anorexia nervosa were interpreted as, at least in part, anxiety-related activation.4 Others used basic taste stimuli, as less complex and less appetitive stimuli may be better suited to identify more basic taste processing as opposed to cognitive and emotional response. In paradigms that applied sugars or aversive taste stimuli, individuals recovered from anorexia nervosa had reduced brain response to repeated, but increased response to randomly applied taste stimuli in the insula, orbitofrontal and striatal regions.5–7 In those studies, the insula, striatum or orbitofrontal cortex distinguished the groups. These regions are central to food reward processing and meal termination.8 Whether neurotransmitter functions are altered in reward processing in individuals with anorexia nervosa has been less explored.
Brain dopamine (DA) circuitry is of particular interest for research on anorexia nervosa as it is involved in the motivation to eat.9 Previous research in individuals with anorexia nervosa during illness and after recovery indicated DA alterations, including increased DA D2 receptor availability,10–13 but we know little about how such alterations may be clinically important or affect behaviour. Primary location of DA neuron activity is measured in the ventral striatum and midbrain, but DA-mediated brain activation is also distributed across the prefrontal cortex, including the insula.14 Within the brain reward circuitry DA has the role of responding to unexpected receipt or omission of rewards, acts as a learning signal and guides us to approach rewarding stimuli, including food.9,15 Thus, the DA system could be involved in altered food approach or food avoidance in individuals with anorexia nervosa. Additionally, DA response in the brain is adaptive to the nutritional state. For instance, animal studies have indicated that food restriction and weight loss were associated with enhanced DA-related reward response,16 suggesting that self-starvation may sensitize dopaminergic brain reward pathways.
During fMRI, we recently applied in individuals with current anorexia nervosa and controls a taste reward learning task that included the association of visual (conditioned) stimuli with sucrose solution as an unconditioned taste stimulus, and after a learning phase individuals could predict the taste stimulus delivered based on the associated conditioned stimulus.17,18 At times this prediction was violated; that is, after the sucrose solution–predicting visual cue no taste followed, or a conditioned visual cue that predicted no taste stimulus was followed by sugar solution. This mismatch of prediction is called prediction error and is known to stimulate DA neurons.9,17 Computational modelling can be used to calculate the prediction error for each trial17 and regress with brain response across all trials to identify brain regions that are sensitive to unexpectancy and could be related to DA function. Results were controlled for medication use and comorbidity, and participants were studied under tight nutritional short-term control. The anorexia nervosa group showed greater activation in the anteroventral striatum, insula and prefrontal cortex.19 Those results suggested that DA-related pathways may be more responsive or sensitive in individuals with anorexia nervosa than controls, which would be consistent with the aforementioned sensitization of reward pathways in relation to underweight.16 This brain-related hyperresponsiveness in the anorexia nervosa group was accompanied by increased sensitivity to rewarding and punishing stimuli (Sensitivity to Reward and Punishment Questionnaire; SPSRQ20) in individuals with anorexia nervosa,19,21 although those behavioural results did not directly correlate with the brain activation.
The present study aimed to identify whether higher prediction error brain reward response or sensitivity using the same paradigm is also present in individuals recovered from anorexia nervosa. To our knowledge, this is the first study using computational model-derived data to investigate brain reward response in recovered individuals. We hypothesized that those who were long-term recovered from anorexia nervosa would have greater prediction error–dependent brain response than healthy control women in the insula and ventral striatum. Greater brain responsiveness after long-term recovery could be a remnant from the illness or potentially a premorbid condition that might predispose a person to altered food reward processing and maybe even to the development of an eating disorder. We also hypothesized that the recovered anorexia nervosa group would show increased sensitivity to salient stimuli on the SPSRQ, suggesting that this is a trait in that population.
Methods
Participants
Through local advertisements, we recruited women recovered from anorexia nervosa and healthy comparison women for participation in our study. Participants were matched for age and education. To be included in the recovered anorexia nervosa group, women had to have a history of restricting-type anorexia nervosa but have a healthy weight (body mass index [BMI] between 18.5 and 24.9, as per the Centers for Disease Control and Prevention), menstrual cycle, exercise regime and food intake for at least 1 year. The Colorado Multiple Institutional Review Board approved the study. All participants provided written informed consent after receiving a complete description of the study.
Behavioural measures
Psychiatric diagnoses, including anorexia nervosa, or absence of any psychiatric disorders in controls were established using the Structured Clinical Interview for DSM-IV diagnoses, and interviews were conducted by a doctoral-level interviewer (G.K.W.F.). All participants completed the following self-assessments: the Drive for Thinness, Bulimia, and Body Dissatisfaction scales from the Eating Disorder Inventory-3;22 the Harm Avoidance and Novelty Seeking subscales from the Temperament and Character Inventory;23 the Spielberger State and Trait Anxiety Inventory;24 the Beck Depression Inventory;25 and the revised SPSRQ.26
To assess response to study solutions as well as general taste sensitivity across groups, participants completed a taste perception test before brain imaging. Each of the participants were presented with a tray of 7 small, unmarked cups that contained distilled water, 5 sucrose solution strengths (Mallinckrodt Chemicals; 2%, 4%, 8%, 16% and 1 M), or artificial saliva (25 mM of potassium chloride, 2 mM of sodium bicarbonate).17 Participants were unaware of the contents of the individual cups and blindly rated the solutions for sweetness and pleasantness on 9-point Likert scales.
Taste conditioning task
We adapted the design used by O’Doherty and colleagues.17 Participants received 3 taste stimuli during fMRI imaging: 1 mol/L sucrose solution (100 trials), no solution (100 trials), and artificial saliva (80 trials). Participants learned to associate each taste stimulus with a paired conditioned visual stimulus (CS) that was probabilistically associated with the unconditioned stimulus (US). The CS shape associated with no solution (null) was followed in 20% of the trials by sucrose solution (unexpected sucrose receipt, positive prediction error condition), and the CS shape for sucrose was followed in 20% of trials by no solution (unexpected sucrose omission, negative prediction error condition). For each participant, the first 10 trials consisted of a fixed CS shape for sucrose followed by the delivery of unexpected sucrose to establish an initial stable association between the CS sucrose shape and the unexpected sucrose taste.17 All other trials were fully randomized without predetermined order. The taste stimuli were applied using a customized programmable syringe pump (J-Kem Scientific) controlled using E-Prime Software (Psychological Software Tools). Individual taste application was triggered by MRI scanner radiofrequency pulse.27 The task duration was 28 minutes.
Image acquisition
Between 7 am and 8 am on the study day, participants ate a standardized breakfast. Functional MRI was performed between 8 am and 9 am. Brain images were acquired using a GE Signa 3 T scanner. We performed T2*-weighted echo-planar imaging (EPI) for blood oxygen–level dependent (BOLD) functional activity under the following parameters: voxel size 3.4 × 3.4 × 2.6 mm, repetition time (TR) 2100 ms, echo time (TE) 30 ms, angle 70°, 30 slices, interleaved acquisition, 2.6 mm slice thickness with a 1.4 mm gap.
Brain imaging data preprocessing
Brain-imaging data were preprocessed and analyzed using SPM8 software running in Matlab R2009b version 7.9.0 software (MathWorks). Data from each participant were realigned to the first volume, normalized to the Montreal Neurological Institute (MNI) template and smoothed with a 6 mm full-width at half-maximum Gaussian kernel. We manually inspected each image sequence, and images with artifacts or movement greater than 1 voxel size were removed.
Data were modelled with a hemodynamic response function, convolved boxcar function, using a general linear model (GLM), including temporal and dispersion derivatives and autoregression. We applied a 128 s high-pass filter to remove low-frequency fluctuation in the BOLD signal. Motion parameters were applied as regressors in the first-level analysis to correct for individual movement.
Computational model analysis
To test temporal difference model-related brain response, we modelled each participant’s individual prediction error signal based on trial sequence.17,28,29 The predicted value (V circumflex) at any time (t) within a trial is calculated as a linear product of weights (wi) and the presence of the CS at time (t) coded in a stimulus representation vector xi(t), where each stimulus (xi) is represented separately at each moment in time:
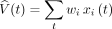
The predicted stimulus value at each time point (t) in the trial is updated by comparing the predicted value at time t + 1 to that actually observed at time t, leading to the prediction error δ(t), where r(t) is the reward at time t:
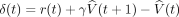
The parameter γ is a discount factor, which determines the extent to which rewards that arrive sooner are more important than rewards that arrive later during the task, with γ = 0.99. The weights (wi) relate to how likely a particular US follows the associated CS, and they are updated on each trial according to the correlation between prediction error and the stimulus representation, where α is a learning rate:
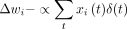
Among various learning rates (i.e., 0.2, 0.5, 0.7) a slow α (0.7) was the best fit for the study groups. The initial reward values were 1 for sucrose and 0 for no solution. The trial-to-trial prediction error was regressed with brain activation across all trials within each participant. For more detailed methods see a previous study (2012) by our group.19
We then regressed the trial-to-trial prediction error with brain activation across all trials within each participant and compared the findings across the groups in a second-level random effects analysis.
Group × condition analysis
In addition to the primary analysis using the computational model, we also conducted an analysis involving first-level models in which we predicted the response in each voxel as a function of each of the 5 stimulus conditions (i.e., expected sucrose, unexpected sucrose, expected no solution, unexpected no solution, expected artificial saliva). Three contrasts of interest were computed: 1) the unexpected sucrose–expected no solution contrast in which the visual stimulus (no solution shape) was the same, but in which there was a positive prediction error signal at the time of the US; 2) the unexpected no solution–expected sucrose contrast in which the visual stimulus was the same, but in which there was a negative prediction error signal at the time of the US; and 3) a contrast across conditioned stimuli comparing the CS that was associated with sucrose with the CS that was associated with no solution. This analysis involved selected trials based on expectation and associated stimulus receipt, but did not take the calculated prediction error into account.
Statistical analysis
For the brain image second-level group comparison, we analyzed either the computational model images or the group × condition first-level contrast images in a GLM whole-brain analysis using SPM8 software. We analyzed a factorial design modelled with the diagnosis as a 2-level factor (control v. recovered anorexia nervosa), with medication use (selective serotonin reuptake inhibitors [SSRIs]) and comorbid depression or anxiety as covariates. We also performed a voxel-wise F test (p < 0.001 uncorrected, extent threshold > 10 voxels). Any significant results at that threshold were then small volume–corrected using the SPM8 automated anatomic labelling (AAL) atlas–derived anatomic regions, family-wise error (FWE)–corrected at p < 0.05.
We analyzed the demographic characteristics and extracted regional brain data using SPSS software version 22 (IBM) and an independent-samples t test.
Results
Participants
Our study included 48 women: 24 recovered from anorexia nervosa and 24 healthy controls. The demographic and clinical characteristics of participants are summarized in Table 1. There were no significant differences in mean age or BMI between the groups. Among the recovered women, the duration of illness with anorexia nervosa ranged between 1 and 21.5 years. All participants were right-handed and had no history of head trauma, neurologic disease, major medical illness, psychosis, or substance use disorders. Thirteen women in the control group and 7 in the recovered anorexia nervosa group took contraceptive pills. Five of the recovered women were taking SSRIs. None of the participants were taking antipsychotics or other psychoactive medications. Among the recovered women, 3 had a mood disorder, 4 had an anxiety disorder (2 specific phobia and 2 with generalized anxiety disorder [GAD]), and 2 had comorbid mood and anxiety disorders (1 with obsessive–compulsive disorder [OCD], GAD and dysthymia, and 1 with panic disorder, GAD and major depressive disorder [MDD]). No woman recovered from anorexia nervosa had a substance use disorder.
Demographic and behavioural characteristics for controls and women recovered from anorexia nervosa
Behavioural results
The participants’ behavioural data are also summarized in Table 1. Women recovered from anorexia nervosa showed greater harm avoidance, depression, drive for thinness, body dissatisfaction and state and trait anxiety than controls. Sensitivity to punishment (SPSRQ) was significantly higher in recovered women than controls. Study groups rated pleasantness and sweetness of study taste solutions similarly.
Brain imaging results
Computational model analysis
Within both controls and recovered women, as expected there was bilateral insula, striatum and midbrain activation in the prediction error whole brain regression. Between groups, the GLM with medication and comorbid mood and anxiety scores as covariates indicated greater brain activation in recovered women than in controls in the left posterior insula (x, y, z = −40, −18, 12; 42 voxels) and right premotor area (x, y, z = 32, −10, 64; 12 voxels). However, only the insula region was significant after small-volume correction (pFWE < 0.05). No regions were significantly more activated in controls than recovered women (Fig. 1). Figure 2 illustrates brain activation in controls compared with recovered women broken down by whether women recovered from anorexia nervosa had comorbid depression, anxiety, or both, and which of those individuals were taking SSRIs.

(A) Increased activation in the left posterior insula in women recovered from anorexia nervosa (AN) compared with controls (family-wise error < 0.05 small-volume correction). (B) Extracted parameter estimates (PEs) across groups.

Brain activation in the left posterior insula, including comorbidity and medication use. White diamonds indicate use of selective serotonin reuptake inhibitors. NO = no comorbid diagnoses; PE = parameter estimate; REC AN = women recovered from anorexia nervosa.
The extracted activation for the left posterior insula did not correlate with any behavioural or demographic values, including duration of illness or recovery.
Group × condition analysis
The unexpected sucrose–expected no solution contrast did not show a significant difference between the groups. Conversely, the unexpected no solution–expected sucrose contrast showed a significant group difference in the left posterior insula (x, y, z = −42, −14, 12); the cluster extent was 15 voxels, but the FWE correction did not show a significant result (p = 0.09). There was no significant group difference for the conditioned stimuli contrast (comparing the sucrose CS with the no solution CS).
Discussion
To our knowledge, this is the first study that investigated computational model-based reward learning in women with anorexia nervosa after recovery. Our primary hypothesis was that recovered women would show enhanced brain reward prediction error response relative to controls. In keeping with our prediction, the results indicate that, compared with controls, women recovered from anorexia nervosa had significantly greater brain activation in the left posterior insula, but no other region. This finding is in contrast to our previous results in ill, underweight individuals with anorexia nervosa. Also notable, SPSRQ sensitivity to punishment but not reward was greater in women recovered from anorexia nervosa than controls.
Our previous data from ill, underweight individuals with anorexia nervosa also showed increased brain activation in this task, but in more regions and in regions that are more typically associated with DA function, such as the ventral striatum.19 The present results in women recovered from anorexia nervosa suggest that those alterations may recover with weight restoration, but that increased response in the posterior insula may be a biological marker long into the recovered state. Alternatively, a tendency toward heightened response in the posterior insula could be a trait, and with weight loss other regions get sensitized. Our previous study that compared individuals with anorexia nervosa, obese individuals and controls found increased activation in the anorexia nervosa group in the anterior insula, and we are currently conducting a larger follow-up study using a design with greater statistical power in order to test whether we find posterior insula activation in individuals currently ill with anorexia nervosa. The group × condition analysis suggested that the group difference may have been driven by the negative prediction error response, but this needs further clarification. Although there is inconsistency with the definition of recovery,30 our sample included only participants who did not meet anorexia nervosa criteria for at least 1 year, and all participants had a BMI of 20 or above except for 1 woman with a BMI of 18 and 1 with a BMI of 19. Therefore, we think that this sample represents a well-recovered group. However, despite their long-term weight recovery and not meeting criteria for anorexia nervosa anymore, the recovered anorexia nervosa group scored significantly higher than controls on drive for thinness and body dissatisfaction. This finding is comparable to those of other studies after long-term recovery31,32 and indicates that cognitive emotional recovery may take much longer than recovery from behaviours that constitute the diagnosis. The neurobiological mechanisms that underlie those behaviours have not been well established.
The posterior insula has been associated primarily with sensorimotor integration and, in general, interoception of bodily feelings,33–35 although recent data suggest that the posterior insula may also entail a “hedonic hotspot” that helps regulate taste pleasantness.36 Importantly, the insula is a central brain region to track prediction error and unexpectancy. Typically, the anterior insula has been associated with that function, including risk prediction,37 but the posterior insula tracks changes of interoceptive states and somatosensory processes.38 This is of interest and importance for anorexia nervosa research because of the disorder’s (ill or recovered) association with distorted body perception that has been linked to the insula among other brain regions,39,40 reduced pain perception that has been linked to the posterior insula,41,42 reduced interoception measured by heart rate detection tasks,43 and the altered drive to eat. Thus, altered activation in the posterior insula could be a potentially important marker for altered pathophysiology in individuals recovered from anorexia nervosa; however, this needs further study.
What could heightened response sensitivity in women recovered from anorexia nervosa mean for behaviour? We did not find direct correlations between posterior insula activation and measured behaviours; however, women recovered from anorexia nervosa had increased sensitivity to salient stimuli, and here especially sensitivity to punishment. It seems, though, that sensitivity to salient stimuli improves with recovery from anorexia nervosa. That is, while individuals currently ill with anorexia nervosa showed increased sensitivity to both reward and punishment,21 we found higher sensitivity to punishment only in the recovered anorexia nervosa group. Mean intolerance of uncertainty was greater in the recovered anorexia nervosa group than the control group, but not to the extent seen in individuals currently ill with anorexia nervosa (p = 0.10).44 Based on those results we suggest that when ill, brain function in individuals with anorexia nervosa is highly sensitive to unexpectancy as well as uncertainty and any type of salient stimulus, rewarding or punishing.21 However, this brain response as well as associated behaviours can improve greatly with weight restoration and recovery. Still, even after long-term recovery posterior insula activation is elevated compared with controls, as is sensitivity to punishment. Those results could be remnants from the illness, but could also be traits; with weight loss and sensitization of DA neurons,45 other areas, including the basal ganglia, may start to show hyperresponsiveness.12
Although greater activity in the posterior insula may indicate heightened sensitivity to the prediction error task, we cannot exclude the possibility that the increased activation may also be linked to emotions, such as anxiety or depression. At the neuronal level, amplified insula response to emotionally salient stimuli has been observed in individuals with anxiety and depression,46 and caloric stimuli typically induce anxiety in individuals with anorexia nervosa. We also found the typical group differences of elevated subjective ratings of anxiety and depression in women recovered from anorexia nervosa compared with controls, which could be further in line with this point. We mitigated those effects by controlling for comorbid diagnosis to capture clinically significant levels of anxiety or depression. Anxiety or depression self-assessment ratings were not correlated with brain response.
This study raises several important points. First, it further highlights that the insula could be a central region of psychopathology in individuals with anorexia nervosa. Previous studies found functional and structural insula alterations in individuals with anorexia nervosa after recovery.6,46–48 Previously, the anterior part of the insula has mostly been implicated in those studies, and it had been hypothesized whether anterior insula alterations could interfere with normal interoceptive awareness and cognitive control circuits.49 The present study, using a sophisticated computational reward learning design and rigorous analysis methods, suggests that the posterior insula could be altered in individuals with anorexia nervosa.
Furthermore, these data raise clinically relevant questions with regard to how symptoms in individuals with eating disorders may be related to erroneous interoceptive tracking or feedback. Prior research indicates that anorexia nervosa is associated with impairments across a variety of sensory stimuli (tactile, haptic, proprioceptive)50 and with increased detection of interoceptive sensations in a meal situation.51 With respect to treatment, understanding body signals may be a key intervention to help reduce anxiety in individuals with anorexia nervosa, and even the knowledge of biological correlates that underlie bodily sensations can be helpful to better tolerate those feelings.
Another implication of brain function and structure results specific to individuals with anorexia nervosa is that those results could be used to build models of brain function.52,53 A problem, though, might be the typically different methods used across studies of eating disorders specifically54 and psychiatry research in general, and it is probably still too early to develop more comprehensive models.55 However, building on the aforementioned publications,52,53 our results in recent years using similar methods suggest larger orbitofrontal cortex volume in individuals with anorexia nervosa across ages and states of illness, as well as greater insula response to the prediction error model task.
Limitations
Our study has some limitations that must be considered. The sample size was not large, although at least 20 participants per cell are usually regarded as providing high reliability.56 A potentially confounding factor was the effect of duration of illness on brain function. The recovered anorexia nervosa group included individuals who had been ill for up to 15 years before recovery. Duration of illness or recovery did not predict brain response. However, the exact number of months or years sick and relapse rates are difficult to reliably quantify. A prospective study including quantifiable measures of duration of illness may be beneficial in future. The effects from comorbid conditions cannot be ruled out; however, we controlled for those variables. In addition, all participants were studied during the same menstrual cycle phase and after eating a standard breakfast. The prediction error task has been associated with DA function, but the method used in our study cannot directly measure any indices of DA neuron activation, and neurotransmitter studies or studies using DA challenge medications will be needed to become more specific in terms of underlying neurotransmitter alterations.
Conclusion
This study involving a prediction error model during brain imaging suggests that anorexia nervosa after recovery is associated with heightened response in the posterior insula, an area that processes somatosensory information and integrates bodily feelings. This heightened sensitivity could contribute to individuals recovered from anorexia nervosa being easily overwhelmed by internal stimuli, such as body sensations, during eating. Whether this is a premorbid condition or a long-lasting effect from the illness will require further study. However, altered posterior insula activity could be an important neurobiological correlate to further study and test whether it relates to altered body perception in individuals with anorexia nervosa.
Acknowledgements
This work was supported by NIMH grant K23 MH080135-01A2, NIMH grant R01 MH096777, NIMH grant R01MH103436 and by the Davis Foundation Award of the Klarman Family Foundation Grants Program in Eating Disorders (all GKWF). We would also like to thank all individuals who participated in the study and the staff at the Eating Disorders Center Denver.
Footnotes
Competing interests: R. O’Reilly is the Chief Scientific Officer at eCortex Inc. No other competing interests declared.
Contributors: G. Frank, M. Shott and R. O’Reilly designed the study. G. Frank and M. Shott acquired and analyzed the data, which S. Collier also analyzed. G. Frank, S. Collier and M. Shott wrote the article, which all authors reviewed and approved for publication.
- Received March 28, 2015.
- Revision received July 10, 2015.
- Accepted September 4, 2015.
References
In this issue

{kind=link}
{kind=link}
Article tools






Related Articles
Cited By...
- No citing articles found.