
Abstract
Asymmetric frontal brain activity has been widely implicated in reactions to emotional stimuli and is thought to reflect individual differences in approach–withdrawal motivation. Here, we investigate whether asymmetric frontal activity, as a measure of approach–withdrawal motivation, also predicts charitable donations after a charity’s (emotion-eliciting) promotional video showing a child in need is viewed, in a sample of 47 young adult women. In addition, we explore possibilities for mediation and moderation, by asymmetric frontal activity, of the effects of intranasally administered oxytocin and parental love withdrawal on charitable donations. Greater relative left frontal activity was related to larger donations. In addition, we found evidence of moderation: Low levels of parental love withdrawal predicted larger donations in the oxytocin condition for participants showing greater relative right frontal activity. We suggest that when approach motivation is high (reflected in greater relative left frontal activity), individuals are generally inclined to take action upon seeing someone in need and, thus, to donate money to actively help out. Only when approach motivation is low (reflected in less relative left/greater relative right activity) do empathic concerns affected by oxytocin and experiences of love withdrawal play an important part in deciding about donations.
Similar content being viewed by others

Donating to charity is a uniquely human form of prosocial behavior that does not directly benefit the individual who donates. As such, theories explaining charitable donating have focused on indirect or psychological benefits, such as establishing a favorable reputation (Nowark & Sigmund, 2005) or the “warm glow” people experience from doing something good or righting a perceived injustice (Andreoni, 1990; Mayo & Tinsley, 2009). Empathic concern for those in need of help may be an especially important motivator for donating to charity, predicting donations even when sociodemographic variables (age, income, and gender) and past donating behavior are taken into account (Verhaert & Van den Poel, 2011).
Various factors may contribute to (the development of) prosocial attitudes and empathy and, thus, affect donating behavior. Recently, we reported on the effects of intranasally administered oxytocin and experienced parental love withdrawal on monetary donations in a donating task, in which participants watched a promotional video asking them to donate some money to UNICEF, after they had just earned 50 Euros by participating in an ERP experiment (Van IJzendoorn, Huffmeijer, Alink, Tops, & Bakermans-Kranenburg, 2011). Oxytocin is a neuropeptide hormone that is increasingly shown to facilitate prosocial behavior (at least in in-group situations; De Dreu et al., 2010). Oxytocin has been found to attenuate stress responses in social situations, to influence the processing of and facilitate memory for salient social stimuli, to promote trust and generosity toward an opponent (for reviews, see Heinrichs, von Dawans, & Domes, 2009; MacDonald & MacDonald, 2010), and to increase the amount of money donated to charity (Barraza, McCullough, Ahmadi, & Zak, 2011).
However, we found that oxytocin increased donations only for those reporting lower parental use of love withdrawal. Love withdrawal is a disciplinary strategy that involves withholding signals of love and affection when a child misbehaves or fails at a task (e.g., ignoring the child, telling the child how sad he/she makes you). When used excessively, it is considered psychological maltreatment (Euser, Van IJzendoorn, Prinzie, & Bakermans-Kranenburg, 2010). By using love withdrawal, the parent communicates to the child that his or her love and affection for the child are conditional upon the child’s compliance and success. The formation of this link between compliance or performance, on the one hand, and relational consequences, on the other, is thought to underlie both the effectiveness and emotional costs of love withdrawal (Assor, Roth, & Deci, 2004; Elliot & Thrash, 2004). Parental use of love withdrawal has been associated with fear of failure, low self-esteem, low emotional well-being, and feelings of rejection and resentment toward the parents in adolescence and young adulthood (Assor et al., 2004; Elliot & Thrash, 2004; Goldstein & Heaven, 2000; Renk, McKinney, Klein, & Oliveros, 2006; Soenens, Vansteenkiste, Luyten, Duriez, & Goossens, 2005). This may bias decision making in social situations away from other-oriented (e.g., empathy for a child in need) to self-oriented (doing what relevant others expect, out of fear for negative reactions) concerns and may hinder empathic concern, leading to lower levels of prosocial behavior (Kanat-Maymon & Assor, 2010; Koenig, Cicchetti, & Rogosh, 2004). Effects of oxytocin may critically depend on individual characteristics and (social) experiences (Bartz, Zaki, Bolger, & Ochsner, 2011). Accordingly, recent studies have found effects of oxytocin to be less pronounced in individuals with (relatively) unfavorable childhood relationship experiences (Bakermans-Kranenburg, Van IJzendoorn, Riem, Tops, & Alink, in press ; Meinlschmidt & Heim, 2007).
To extend our previous findings, we now turn our attention to another characteristic that has been implicated in emotional and motivational processes: asymmetric frontal cortical activity. Differences in power within the alpha band (8–12 Hz) of the electroencephalogram (EEG) over left and right frontal areas are widely used to quantify asymmetric frontal brain activity. Because greater alpha power is related to deactivation of the underlying cortical tissue (Cook, O’Hara, Uijtdehaage, Mandelkern, & Leuchtner, 1998; Laufs et al., 2003), greater alpha power over right than over left frontal areas reflects greater activity of the left than of the right frontal cortex, whereas greater alpha power over left than over right frontal areas reflects relatively greater activity of the right frontal cortex. Numerous studies have related asymmetric frontal activity to emotional processes in individuals of all ages (e.g., Coan & Allen, 2004; Davidson & Fox, 1989; Fox, Henderson, Rubin, Calkins, & Schmidt, 2001). Whereas early studies focused on emotional valence, showing a relation between greater relative left activity and a tendency to experience certain positive emotions (e.g., happiness) and between greater relative right activity and a tendency to experience certain negative emotions (e.g., fear), more recent research has suggested that asymmetric frontal activity relates to motivational direction (of emotions), rather than to emotional valence (for a review, see Harmon-Jones, Gable, & Peterson, 2010). Frontal asymmetries seem to be best characterized as reflecting a general tendency for approach versus withdrawal, with greater left activity reflecting greater approach motivation and greater right activity reflecting greater withdrawal motivation, although there is more evidence for the link between left frontal activity and approach than for the link between right frontal activity and withdrawal (Demaree, Everhart, Youngstrom, & Harrison, 2005; Harmon-Jones & Allen, 1997; Harmon-Jones et al., 2010). Measures of asymmetric frontal activity have been shown to track both a general trait of and state-related fluctuations in approach–withdrawal motivation, with the contributions of trait- and state-related variation estimated to be about 50% each (Coan & Allen, 2004; Hagemann, Hewig, Seifert, Naumann, & Bartussek, 2005).
Although little research into the neural origins of asymmetric electrocortical activity has been conducted, the available evidence suggests that electrocortical measures of frontal asymmetry reflect activity in (dorso)lateral prefrontal areas (Pizzagalli, Sherwood, Henriques, & Davidson, 2005; see also Davidson, 2004). Evidence from animal studies suggests that the dorsolateral prefrontal cortex is involved in the processing of reward-related information in goal-directed behavior (Kobayashi, Lauwereyns, Koizumi, Sakagami, & Hikosaka, 2002; Wallis & Miller, 2003). Studies in humans have shown differential effects of positively and negatively valenced stimuli both on left and right (dorso)lateral prefrontal brain activity and on working memory performance, pointing toward the integration of cognitive and emotional material in this brain region (Gray, Braver, & Raichle, 2002; Herrington et al., 2005). Although much remains to be investigated, both lines of evidence are consistent with a role of lateral prefrontal areas in approach–withdrawal motivation (see also Davidson, 2004).
As a measure of approach–withdrawal motivation, asymmetric frontal activity may be expected to relate to donating behavior when individuals are confronted with a charity’s promotional material (typically showing the precarious situation of those in need) and are asked for a donation. Higher approach motivation and a greater tendency to experience approach-related emotions, associated with greater relative left frontal activity, may well cause an individual to donate more money to actively help out those in need. Thus, we expect that asymmetric frontal activity will predict monetary donations to UNICEF in the context of our donating task.
Because asymmetric frontal activity both reflects a trait-like motivational tendency to react in a certain (predictable) way in emotionally evocative situations and is susceptible to state-related changes, it has been suggested that frontal asymmetries may serve as moderators or mediators of behavior (Coan & Allen, 2004). Therefore, we will also investigate whether frontal alpha asymmetry mediates or moderates the effect of oxytocin administration and parental love withdrawal that we have found previously (Van IJzendoorn et al., 2011).
For mediation to occur, asymmetric frontal activity should not only be related to donating behavior, but also be affected by oxytocin, parental love withdrawal, or both. Many effects of oxytocin on socio-emotional information processing (e.g., increased processing of facial expressions; Domes et al., 2010; Huffmeijer, Alink, et al., 2011) and behavior (e.g., increases in trust and generosity; Zak, Kurzban, & Matzner, 2005; Zak, Stanton, & Ahmadi, 2007) may be linked to approach motivation, suggesting that oxytocin might increase relative left frontal activity. Also, oxytocin has been found to have anxiolytic effects in social situations (Heinrichs, Baumgartner, Kirschbaum, & Ehlert, 2003), and anxiety has been related to increased relative right frontal activity (e.g., Thibodeau, Jorgensen, & Kim, 2006). In fact, it was recently argued that increasing approach motivation may be one of the mechanisms through which oxytocin achieves its prosocial effects (Kemp & Guastella, 2011).
Experiences of parental love withdrawal might also relate to frontal alpha asymmetry. Experiences of parental love withdrawal have been related to characteristics associated with social withdrawal (e.g., low emotional well-being and low self-esteem [see, e.g., Assor et al., 2004; Renk et al., 2006]) and fear of failure (i.e., anxiety in performance situations [see, e.g., Elliot & Thrash, 2004; Soenens et al., 2005]). Thus, experiences of love withdrawal may be related to withdrawal tendencies. In addition, measures of frontal alpha asymmetry for the present study were collected before and after participants performed a computerized feedback task (i.e., in a performance situation). Because relative right frontal activity may be related to anxiety primarily within anxiety-provoking situations (Crost, Pauls, & Wacker, 2008), in this context, parental love withdrawal might be related to relative right frontal activity.
In the case of moderation, on the other hand, effects of oxytocin and parental love withdrawal would depend on an individual’s level (direction and degree) of asymmetric frontal activity. Effects of oxytocin and concerns related to experiences of parental love withdrawal might, for example, exert less influence over decision making in the donating task for those showing greater relative left frontal activity, who may be expected to respond positively to a request for a donation after seeing an individual in need, because of a high level of approach motivation.
To summarize, the present study examines whether asymmetric frontal brain activity, as a measure of approach–withdrawal motivation, predicts donations to charity after a promotional video of a child in need is viewed. We expect that greater relative left frontal activity, reflecting higher approach motivation, predicts larger donations. In addition, we explore the possibility that asymmetric frontal brain activity mediates or moderates the combined effect of oxytocin and parental love withdrawal on donating behavior.
Method
Participants
A total of 59 female undergraduate students, 18–30 years of age (M = 20.54, SD = 2.89), participated in the study. Two participants did not complete the donating task (because they did not participate in the second session, in which this task took place), 9 participants contributed insufficient EEG data because of excessive ocular or motion artifacts, and data collection for 1 participant was disturbed by loud noise. The final sample thus consisted of 47 female undergraduate students, 18–30 years of age (M = 20.45, SD = 2.80). Exclusion criteria included color blindness, smoking, alcohol and drug abuse, neurological and psychiatric disorders, pregnancy, breast-feeding, and use of medication (except oral contraceptives). The study was approved by the ethics committee of the Leiden University Medical Center, and informed consent was obtained from participants at the beginning of the experiment.
Procedure
Participants completed a questionnaire measuring parental use of love withdrawal during an introductory course in child and family studies. The questionnaire was administered to 391 18- to 30-year-old female undergraduate students who were willing to participate in an ERP experiment. Participants for this experiment were selected stratified from the pool of 391 students, on the basis of their scores on the maternal version of the questionnaire: Half of the participants were selected randomly from the group scoring in the upper quartile of the questionnaire (n = 23 for the present sample), and half of the participants were selected randomly from the group scoring in the other three quartiles (n = 24 for the present sample), resulting in a normal distribution of love withdrawal scores (see below).
Participants were asked to come to our laboratory for two experimental sessions, separated by approximately 4 weeks. To minimize influences of diurnal variations in oxytocin levels, all sessions took place in the afternoon (starting between 12:00 and 15:00). Here, we report on the second session, which ended with the donating task. Participants were instructed to abstain from alcohol and excessive physical activity during the 24 h before the start of each session and from caffeine on the day of the session.
Concerning the administration of oxytocin, participants were told that they would receive oxytocin during one session and a placebo during the other and that the order was not known even to the experimenter. This message was repeated at the beginning of the second session. Participants were not informed about the effects of oxytocin under investigation, only about the possible side effects they might experience (as required by the ethics committee).
At the start of each session, participants received a nasal spray containing either 15.6 IU of oxytocin or a placebo (saline solution). All participants received both substances once, either the placebo during the first session and oxytocin during the second or oxytocin during the first session and the placebo during the second. The order of administration was unknown to both the participant and the experimenter and was counterbalanced across participants. Thus, during the second session, in which the donating task took place, half of the participants received oxytocin (n = 22), and half a placebo (n = 25). For the present study, the evaluation of effects of oxytocin thus involves a between-subjects comparison. Participants were then fitted with an electrode net, after which their EEG was recorded during two 2-min resting periods (the first with eyes opened, the second with eyes closed). Participants then completed a 1-h task (for ERP data collection; results presented elsewhere), after which their EEG was again recorded during two 2-min resting periods (again, the first with eyes opened, the second with eyes closed).
Donating to UNICEF
After completion of the last EEG measurements of the second session and after removal of the electrode net, participants were paid 50 Euros for participation. They were then left alone and were shown a 2-min UNICEF promotional video, showing a child from a resource-limited country (Bangladesh), forced to work in a stone pit instead of going to school, due to poverty. Immediately following the video, a text appeared on screen, asking the participant to donate some money. A money box had been positioned next to the video screen. The money box was filled with several coins to enhance credibility (see Van IJzendoorn, Bakermans-Kranenburg, Pannebakker, & Out, 2010, for a similar task). Because the distribution of donations was somewhat skewed, a square root transformation was used in all analyses. Donated money was transferred to the UNICEF bank account after data collection.
EEG measurement
During EEG data collection, participants were seated in a comfortable chair facing a computer screen (distance approximately 50 cm), in a dimly lit, sound-attenuated room. A white fixation cross was presented on a black background at the center of the screen during resting measurements with eyes opened. Participants were instructed to “just relax” and keep their eyes focused on the cross as much as possible. Participants’ EEG was acquired during the four resting periods, using 129-channel hydrocel geodesic sensor nets, amplified using a NetAmps300 amplifier, low-pass filtered at half (i.e., 125 Hz) the digitization rate of 250 Hz, and recorded using NetStation software (Electrical Geodesics, Inc.). Impedances were kept below 50 kΩ. Further processing of the raw EEG was conducted offline using Brain Vision Analyzer (BVA) 2.0 software (Brain Products). The EEG was filtered with a passband range of 0.1–40 Hz (−3 dB, 48 dB/octave) and rereferenced to the average of activity in all channels. Each 2-min recording was divided into one hundred nineteen 2-s segments, with 1-s overlap between segments, and was corrected for ocular artifacts using Independent Components Analysis: The algorithm implemented in BVA 2.0 was set to automatically select those components for deletion that explained the largest amount of relative variance in the vertical EOG (electrodes 25–127; several components, together explaining up to 15% of relative variance, could be selected if the first component explained less than 15%) and horizontal EOG (electrodes 128–125; several components, together explaining up to 10% of relative variance, could be selected if the first component explained less than 10%). After visual inspection of the scalp distributions of the components’ activity, the automatic selection was corrected if necessary: Automatically selected components were unmarked for deletion if they did not show the typical distribution of either blinks and vertical eye movements (positive field above and negative field below the eyes or vice versa) or horizontal eye movements (positive field at one eye and negative field at the other). Additional components (maximum, 3) were marked for deletion if they showed a distribution typical for blinks or eye movements. The algorithm then proceeded with the actual data correction.
Segments containing residual artifacts were removed (segments were removed if the slope at any point during the segment exceeded 100 μV/ms and if the difference between the maximum and minimum activity exceeded 300 μV within the entire segment or was less then 0.5 μV within any 100-ms period), and a short term Fourier transform (0.5-Hz resolution, 100% Hamming window) was computed to obtain power values (μV2) for the remaining segments. Power values were averaged across all segments within each resting period and then were averaged across the frequency range of 8–12 Hz to obtain measures of power within the alpha band within each resting period. To normalize data distribution, the natural logarithm (ln) of these values was computed.
Ln-transformed values were averaged across sets of eight electrodes to yield measures of left-frontal (20, 23, 24 [F3], 26, 27, 28, 33 [F7], 34), right-frontal (2, 3, 116, 117, 118, 122 [F8], 123, 124 [F4]), left-central (30, 36 [C3], 37, 40, 41, 42, 45, 46), right-central (87, 93, 102, 103, 104 [C4], 105, 108, 109), left-posterior (52 [P3], 53, 58 [P7], 59, 60, 61, 64, 65), and right-posterior (78, 85, 86, 90, 91, 92 [P4], 95, 96 [P8]) alpha activity. As a measure of asymmetry, laterality indices were computed separately for frontal, central, and posterior sites by subtracting left alpha activity from right alpha activity (ln[right alpha] – ln[left alpha]). A zero value on this measure thus represents no alpha asymmetry, whereas more positive values result from greater alpha power over right than over left cortical areas and, therefore, represent greater relative left cortical activity. Similarly, more negative values represent greater relative right cortical activity. Because no effects involving time (resting periods before vs. after the ERP experiment) or condition (eyes open vs. closed) were found (all Fs ≤ 2.24, ps > .10) in an ANOVA with caudality (frontal, central, posterior), drug (placebo vs. oxytocin), time, and condition as independent variables, asymmetry values were averaged across the four baseline periods for statistical analyses. Cronbach’s alpha for asymmetry values, with laterality indices for individual electrode pairs (e.g., F4–F3) as items, was .85 for frontal, .82 for central, and .84 for posterior asymmetry. Frontal asymmetry values were modestly stable across the two experimental sessions (r = .47, p < .01), consistent with contributions of both state-related and trait-related variation.
Parental use of love withdrawal
Participants completed an 11-item questionnaire, containing all 5 items of the Withdrawal of Relations subscale of the Children’s Report of Parental Behavior Inventory (Beyers & Goossens, 2003; Schludermann & Schludermann, 1988), 2 items that were adapted from this same questionnaire, and 4 items adapted from the Parental Discipline Questionnaire (Hoffman & Saltzstein, 1967; Patrick & Gibbs, 2007). Participants rated how well each of the 11 statements described their mother and father separately (e.g., “My mother is a person who, when I disappoint her, tells me how sad I make her”) on a 5-point scale ranging from 1 (not at all) to 5 (very well). Scores for maternal and paternal love withdrawal were summed, and the resulting scale was normally distributed. Cronbach’s alpha was .87 for the present sample.
Analyses
Statistical analyses were performed using SPSS 17 software. To evaluate effects (simple, mediation, moderation) of frontal alpha asymmetry on donating behavior, a series of hierarchical linear regression analyses were performed. In all regression analyses, continuous predictors were centered on their respective means, and contrast codes were used for drug (placebo vs. oxytocin).
Results
Descriptives
Among the 47 participants included in the present sample, 37 (80%) donated some money (range, 0.50–15.00 Euros), and the average amount of money donated was 2.77 Euros (SD = 2.83). As described above, a square root transformation was computed to normalize data distribution, and transformed values were used in all analyses. The average score on the love withdrawal questionnaire was 48.06 (SD = 12.75); average values for frontal, central, and posterior alpha asymmetry were −0.05 (SD = 0.17), −0.10 (SD = 0.26), and 0.07 (SD = 0.27), respectively. Posterior alpha asymmetry was significantly correlated with both frontal (r = −.45, p < .01) and central (r = .39, p < .01) alpha asymmetry, whereas frontal and central asymmetry were not significantly related (r = .16, p > .10).
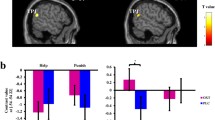
To confirm our previous result that the interaction between drug and love withdrawal predicted donating behavior in the present sample of 47 students, a hierarchical regression analysis was performed with drug (placebo vs. oxytocin) entered in the first step, love withdrawal in the second step, and the interaction between drug and love withdrawal in the third step. As was expected, the model was significant, F(3, 43) = 2.91, p < .05, R 2 = .17, and only the interaction term significantly predicted donating (β = .32, p < .05; other |βs| ≤ .16, ps > .10). As can be seen in Fig. 1, oxytocin increased donations only for those reporting lower love withdrawal.

Mean donations (square root transformation) after placebo and oxytocin administration for participants reporting low (n = 24) and high (n = 23) love withdrawal. Participants reporting lower love withdrawal donated more money to a charity (UNICEF) after oxytocin than after placebo administration. Participants were divided into groups for display purposes only
Frontal alpha asymmetry and donating
Simple effect
Frontal alpha asymmetry and donating behavior were significantly and positively correlated (r = .30, p < .05). More positive values and, thus, greater relative left frontal activity were related to larger donations to UNICEF (illustrated in Fig. 2).

Scatterplot and estimated regression line illustrating the relation between asymmetric frontal cortical activity and donations to UNICEF. Greater relative left frontal activity (more positive asymmetry values) was associated with larger donations
Mediation
To test whether frontal alpha asymmetry mediated effects of oxytocin and love withdrawal on donating behavior, we first examined whether frontal alpha asymmetry was predicted by oxytocin, love withdrawal, or both. We therefore conducted a hierarchical regression analysis with frontal alpha asymmetry as the dependent variable, in which drug (placebo vs. oxytocin) was entered as a predictor in the first step, love withdrawal in the second step, and the interaction term in the third step. No significant effects were found [all |βs| ≤ .10, ps > .50; model: F(3, 43) = 0.15, p > .50 R 2 = .01], thus ruling out mediation.
Moderation
To test for potential moderating effects of frontal alpha asymmetry, a hierarchical regression analysis was performed with donation as the dependent variable, in which drug (placebo vs. oxytocin) was entered in the first step, love withdrawal and frontal alpha asymmetry in the second step, all two-way interactions of the (centered) variables (drug × love withdrawal, drug × frontal alpha asymmetry, love withdrawal × frontal alpha asymmetry) in the third step, and the three-way interaction (drug × love withdrawal × frontal alpha asymmetry) in the fourth step. The final model was significant, F(7, 39) = 3.71, p < .01, R 2 = .40. As in the analyses described above, the main effect of frontal alpha asymmetry (β = .30, p < .05) and the interaction between drug and love withdrawal (β = .27, p < .05) were significant. These effects were, however, qualified by a significant three-way interaction between drug, love withdrawal, and frontal alpha asymmetry (β = −.32, p < .05). No other effects were significant (all |βs| ≤ .22, ps > .10). To further explore the three-way interaction, we divided the participants into groups showing above-average (more positive values, reflecting greater relative left/less relative right activity, n = 23) and below-average (more negative values, reflecting greater relative right/less relative left activity, n = 24) frontal alpha asymmetry and then conducted separate hierarchical regression analyses for each group, with donation as the dependent variable, and predictors drug (placebo vs. oxytocin) entered in the first step, love withdrawal in the second, and the interaction term in the third step. For the group showing greater relative right/less relative left activity, the model was significant, F(3, 20) = 8.40, p < .01, R 2 = .56, and the interaction between drug and love withdrawal significantly predicted donations (β = .68, p < .01; other |βs| ≤ .24, ps > .10), whereas for the group showing greater relative left/less relative right activity, no significant effects were found [all |βs| ≤ .32, ps > .10; model: F(3, 19) = 0.91, p > .10, R 2 = .13]. As can be seen in Fig. 3, lower love withdrawal was associated with larger donations after oxytocin (as compared with placebo) administration for those showing greater relative right frontal activity.

Differences between donations (square root transformation) to a charity after oxytocin versus placebo administration (predicted donation after oxytocin – predicted donation after placebo) as a function of frontal alpha asymmetry and love withdrawal. The two regression lines illustrate the relation between love withdrawal and the effect of oxytocin on donations for relatively positive (greater relative left frontal activity; gray line) and relatively negative (greater relative right frontal activity; black line) asymmetry values. For those showing greater relative right frontal activity (more negative asymmetry values; illustrated in black), lower love withdrawal was associated with larger donations in the oxytocin than in the placebo condition. * Mean LW: mean score on the love withdrawal questionnaire across participants. Mean LW − 1 SD represents relatively low love withdrawal; mean LW + 1 SD represents relatively high love withdrawal. ** Mean asymmetry: mean frontal alpha asymmetry score across participants. Note that mean asymmetry − 1SD reflects relative right frontal activity (M − 1SD = −0.05 − 0.17 = −0.22), whereas mean asymmetry + 1 SD reflects relative left frontal activity (M + 1SD = −0.05 + 0.17 = 0.12)
Accounting for central and posterior alpha asymmetry
Because we found some significant correlations between alpha asymmetry at frontal, central, and posterior electrode sites, we repeated the analyses (with donation as a dependent variable) described above under "Simple effect" and "Moderation" twice, once with central and once with posterior alpha asymmetry replacing frontal alpha asymmetry. Neither central (r = .17, p > .10) nor posterior (r = −.06, p > .50) alpha asymmetry was significantly correlated with donations, and in regression analyses, no significant effects involving central (all |βs| ≤ .19, ps > .10) or posterior (all |βs| ≤ .15, ps > .10) alpha asymmetry were found. Finally, we performed a regression analysis (with donation as the dependent variable) in which central and posterior alpha asymmetry were entered as predictors in the first step to control for potential effects of central and posterior alpha asymmetry on donating behavior, before entering drug (placebo vs. oxytocin; step 2), love withdrawal and frontal alpha asymmetry (step 3), and the two- (step 4) and three-way (step 5) interaction terms of these three variables. The final model was significant, F(9, 37) = 2.78, p < .05, R 2 = .40, and both the main effect of frontal alpha asymmetry (β = .33, p < .05) and the interaction between drug, love withdrawal, and frontal alpha asymmetry (β = −.31, p < .01) remained significant. Neither central (β = −.02, p > .50), nor posterior (β = .08, p > .50) alpha asymmetry significantly predicted donations. The interaction between drug and love withdrawal just failed to reach significance (β = .27, p = .05), and no other significant effects were found (all |βs| ≤ .22, ps > .10).
Discussion
Asymmetric frontal brain activity significantly predicted donating behavior. As was expected, greater relative left frontal activity/less relative right frontal activity was associated with larger donations to UNICEF after a promotional video showing a child in need. This finding is in line with the idea that higher approach motivation and a greater tendency to experience approach-related emotions, associated with greater relative left frontal activity, would cause an individual to donate more money to actively help out those in need.
Moreover, frontal alpha asymmetry moderated the interactive effect of oxytocin and parental love withdrawal on donating behavior that we had observed previously (Van IJzendoorn et al., 2011). The predictive value of the interaction between oxytocin and parental love withdrawal (larger donations after oxytocin than after placebo administration for those reporting lower love withdrawal) increased with decreasing relative left/increasing relative right frontal activity. Lower love withdrawal was associated with larger donations after oxytocin than after placebo administration only for those showing relative right frontal activity. Thus, it seems that the effects of oxytocin on prosocial attitudes and behavior and concerns related to experiences of parental love withdrawal affect decisions about donating only for individuals whose response to emotional material is characterized by withdrawal rather than approach, as suggested by their pattern of frontal brain activity. We tentatively suggest that those showing greater relative left frontal activity are likely to donate money in response to promotional material showing an individual in need, irrespective of how their empathic responding is affected by oxytocin administration or experiences of love withdrawal, because approach-related tendencies motivate them to take action and, thus, to donate money. For those less inclined to donate out of approach motivation (i.e., those showing less relative left/greater relative right frontal activity), empathic and other concerns affected by oxytocin and experiences of love withdrawal may play a more important part in deciding whether and how much money they donate.
Neither love withdrawal nor oxytocin predicted asymmetric frontal cortical activity. Thus, exogenously administered oxytocin and experiences of parental love withdrawal do not seem to directly influence approach–withdrawal motivation. It is possible that the effects of love withdrawal and oxytocin would be limited to social, rather than general, approach–withdrawal motivation. Our measure of asymmetric frontal activity cannot be expected to capture this aspect completely, since EEG data were collected during resting periods in a laboratory environment. Alternatively, oxytocin might relate to approach–withdrawal motivation and asymmetric frontal cortical activity only under some conditions and/or for some individuals. In a recent article by Bartz et al. (2011), it was suggested that oxytocin primarily heightens the salience and processing of emotional stimuli, which may have different consequences—for example, for approach–withdrawal motivation—depending on (the interplay between) contextual and personal characteristics.
The most obvious feature of the interaction between oxytocin and parental love withdrawal, both when differences in asymmetric frontal brain activity are ignored (see Fig. 1; cf. Van IJzendoorn et al., 2011) and when individuals showing less relative left/greater relative right frontal activity are focused on (see Fig. 3), is the increase in donations after oxytocin (vs. placebo) administration for individuals reporting lower love withdrawal. However, a closer look at Figs. 1 and 3 reveals that at the higher end of the love withdrawal continuum, oxytocin actually seems to decrease donations, relative to placebo, for those showing less relative left/greater relative right frontal activity. An interpretation of these seemingly opposing effects of oxytocin is necessarily speculative. Nevertheless, experiences of maternal love withdrawal may be related to various aspects of socio-emotional functioning (Assor et al., 2004; Goldstein & Heaven, 2000; Soenens et al., 2005) and information processing (Huffmeijer, Alink, et al., 2011; Huffmeijer, Tops, Alink, Bakermans-Kranenburg, & Van IJzendoorn, 2011) and, potentially, to alterations in the oxytocin system, since evidence suggests that relationship experiences may be important for shaping this system (Feldman, Gordon, & Zagoory-Sharon, 2010; Heim et al., 2008), which could limit or alter the effects of oxytocin (Bartz et al., 2011).
Importantly, several other lines of evidence also suggest that the positive effects of oxytocin administration may be more pronounced in individuals with relatively favorable childhood relationship experiences. Oxytocin administration reduced cortisol levels in adults who had not experienced any major childhood adversity, but not in adults who endured parental separation early in life (Meinlschmidt & Heim, 2007). We found that oxytocin administration decreased the use of excessive force when infant crying was listened to, but only for those who experienced little harsh discipline in their childhood (Bakermans-Kranenburg et al., in press). In fact, we speculate that lowered oxytocin sensitivity might be the result of epigenetic processes (Van IJzendoorn, Bakermans-Kranenburg, & Ebstein, in press). Parental rejection might lead to elevated levels of methylation and, thus, suppress genetic expression, not only in genetic areas related to the glucocorticoid system (McGowan et al., 2009) or the serotonin system (Van IJzendoorn, Caspers, Bakermans-Kranenburg, Beach, & Philibert, 2010), but also in areas related to the oxytocinergic system, which may, in turn, decrease receptiveness to intranasal oxytocin administration (Van IJzendoorn et al., 2011).
Whatever causes individuals varying in experiences of love withdrawal to differentially respond to oxytocin administration, it is important to note that although individuals showing less relative left/greater relative right frontal activity appear to be more sensitive to influences of oxytocin and parental love withdrawal on (processes involved in) decision making in the donating task, these influences are not necessarily unidirectional. As compared with those showing greater relative left frontal activity, individuals showing greater relative right frontal activity may show both larger increases and smaller increases or even decreases in donations after oxytocin than after placebo administration, depending on their experiences of parental love withdrawal.
There is a striking similarity between this observation and findings concerning the model of differential susceptibility of individuals to environmental influences throughout development, showing that children with certain neurobiological dispositions (as evident from genetic, neuroendocrine, or behavioral measures) both benefit more from a favorable caregiving environment and are more affected by negative caregiving experiences (for an overview, see Ellis, Boyce, Belsky, Bakermans-Kranenburg, & Van IJzendoorn, 2011). The differential susceptibility model has been contrasted with diathesis-stress or dual-risk models that focus on the added negative effects of unfavorable circumstances for some individuals, assuming no differential effects of favorable circumstances (see Ellis et al., 2011). Interestingly, a diathesis-stress model has also been proposed for the role of relative right frontal brain activity in depression (Davidson, 1998).
There are, of course, substantial differences between the differential susceptibility literature and our present study (but see Bakermans-Kranenburg & Van IJzendoorn, 2011, for another study on differential susceptibility and donating to a charity). Nevertheless, a recurrent observation in studies of the interplay between neurobiological characteristics and external factors or experiences is that certain characteristics are associated with a greater sensitivity to (influences of) external factors and experiences and that effects of this heightened sensitivity can go both ways: Depending on the level of the external variable or experiences under consideration, the more susceptible or sensitive individual can be both worse and better off, show both more and less desirable behavior, or, as in our present study, show both similar but larger and opposite effects of oxytocin, as compared with the less sensitive individual. Thus, however associative the link between our present results and those of differential susceptibility theory may be, this type of interaction between neurobiological characteristics, experiences, and external factors deserves attention in future studies.
Since the approach–withdrawal model was devised specifically to explain the role of asymmetric frontal brain activity in emotional processes (see, e.g., Demaree et al., 2005; Harmon-Jones et al., 2010), including affective reactions to emotional film fragments and photographs (e.g., Perry, Bentin, Bartal, Lamm, & Decety, 2010; Tomarken, Davidson, & Henriques, 1990), showing a UNICEF promotional video of a child in need may be an important component of the present experimental setup. Showing such a video activates emotional systems (by eliciting emotional reactions such as empathy; see, e.g., Burt & Strongman, 2004) and may thus be particularly suited to study the influences of individual characteristics (in this case, frontal asymmetries and parental love withdrawal) and substances (in this case, oxytocin) involved in emotional processes on donating behavior.
A factor to consider when interpreting our results is that all our participants were female. We included only women in this study because of the considerable differences between males and females in the oxytocin system (Suske & Gallagher, 2009), because the effects of oxytocin in women are less frequently studied than those in men (Bos, Panksepp, Bluthé, & Van Honk, in press), and because the ERP experiment focused on the effects of maternal use of love withdrawal with daughters (see Huffmeijer, Alink, et al., 2011; Huffmeijer, Tops, et al., 2011). Nevertheless, it would be interesting to study the same processes in men. Furthermore, we measured asymmetric frontal cortical asymmetry before the participants engaged in the donating task. To evaluate potential changes in asymmetric frontal activity while the video is watched and to test potential contributions of these changes to donating behavior, future studies could include measures of asymmetric cortical activity during the donating task. Measures of asymmetric frontal activity collected in an emotional context have been suggested to be particularly useful in predicting emotional behavior (Coan, Allen, & McKnight, 2006). Future studies should also include behavioral or questionnaire measures of approach–withdrawal motivation (such as the BIS/BAS scales; Carver & White, 1994), to test our interpretation of the effects of asymmetric frontal activity on donating behavior in terms of approach–withdrawal motivation. The fact that we did not include such a questionnaire measure in the present study limits the validity of our interpretation. Importantly, though, our results were specific to frontal alpha asymmetry and were not affected by the inclusion of central and posterior alpha asymmetry, increasing confidence in our findings. In addition, it may be interesting and aid interpretation to include subjective evaluations of the UNICEF video (e.g., ratings of valence and arousal) in future experiments. Finally, because measures of frontal alpha asymmetry have been found to be composed of both a trait level of asymmetry and state-induced variation (Coan & Allen, 2004; Hagemann et al., 2005), future studies could include multiple measures (taken on multiple occasions) of frontal alpha asymmetry to evaluate the respective contributions of trait-related and state-related factors.
In conclusion, greater left than right frontal brain activity predicted larger donations after a video of a child in need was viewed. Moreover, asymmetric frontal activity moderated the effects of oxytocin and parental love withdrawal on donating behavior. We suggest that when approach motivation is high (reflected in greater relative left frontal activity), individuals are inclined to take action upon seeing someone in need and, thus, to donate more money to actively help out. Only when approach motivation is low (reflected in less relative left/more relative right activity) do empathic and other concerns affected by oxytocin and experiences of love withdrawal play an important part in deciding about donations. Future research, incorporating direct measures of approach–withdrawal motivation, will be necessary to test this interpretation and to extend findings to a wider population, including men.
References
Andreoni, J. (1990). Impure altruism and donations to public goods: A theory of warm-glow giving. The Economic Journal, 100, 464–477.
Assor, A., Roth, G., & Deci, E. L. (2004). The emotional costs of parents’ conditional regard: A self-determination theory analysis. Journal of Personality, 72, 47–88.
Bakermans-Kranenburg, M. J., & Van IJzendoorn, M. H. (2011). Differential susceptibility to rearing environment depending on dopamine-related genes: New evidence and a meta-analysis. Development and Psychopathology, 23, 39–52.
Bakermans-Kranenburg, M. J., Van IJzendoorn, M. H., Riem, M. M. E., Tops, M., & Alink, L. R. A. (in press). Oxytocin decreases handgrip force in reaction to infant crying in females without harsh parenting experiences. Social Cognitive and Affective Neuroscience.
Barraza, J. A., McCullough, M. E., Ahmadi, S., & Zak, P. J. (2011). Oxytocin infusion increases charitable donations regardless of monetary resources. Hormones and Behavior, 60, 148–151. doi:10.1016/j.yhbeh.2011.04.008
Bartz, J. A., Zaki, J., Bolger, N., & Ochsner, K. N. (2011). Social effects of oxytocin in humans: Context and person matter. Trends in Cognitive Sciences, 15, 301–309.
Beyers, W., & Goossens, L. (2003). Psychological separation and adjustment to university: Moderating effects of gender, age and perceived parenting style. Journal of Adolescent Research, 18, 363–382.
Bos, P. A., Panksepp, J., Bluthé, R. M., & Van Honk, J. (in press). Acute effects of steroid hormones and neuropeptides on human social-emotional behavior: A review of single administration studies. Frontiers in Neuroendocrinology. doi:10.1016/j.yfrne.2011.01.002
Burt, C. D. B., & Strongman, K. (2004). Use of images in charity advertising: Improving donations and compliance rates. International Journal of Organisational Behaviour, 8, 571–580.
Carver, C. S., & White, T. L. (1994). Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: The BIS/BAS scales. Journal of Personality and Social Psychology, 67, 319–333.
Coan, J. A., & Allen, J. J. B. (2004). Frontal EEG asymmetry as a moderator and mediator of emotion. Biological Psychology, 67, 7–49.
Coan, J. A., Allen, J. J. B., & McKnight, P. E. (2006). A capability model of individual differences in frontal EEG asymmetry. Biological Psychology, 72, 198–207.
Cook, I. A., O’Hara, R., Uijtdehaage, S. H. J., Mandelkern, M., & Leuchtner, A. F. (1998). Assessing the accuracy of topographic EEG mapping for determining local brain function. Electroencephalography and Clinical Neurophysiology, 107, 408–414.
Crost, N. W., Pauls, C. A., & Wacker, J. (2008). Defensiveness and anxiety predict frontal EEG asymmetry only in specific situational contexts. Biological Psychology, 78, 43–52.
Davidson, R. J. (1998). Affective style and affective disorders: Perspectives from affective neuroscience. Cognition and Emotion, 12, 307–330.
Davidson, R. J. (2004). What does the prefrontal cortex “do” in affect: Perspectives on frontal EEG asymmetry research. Biological Psychology, 67, 219–233.
Davidson, R. J., & Fox, N. A. (1989). Frontal brain asymmetry predicts infants’ response to maternal separation. Journal of Abnormal Psychology, 98, 127–131.
De Dreu, C. K. W., Greer, L. L., Handgraaf, M. J. J., Shalvi, S., Van Kleef, G. A., Baas, M., & Feith, S. W. W. (2010). The neuropeptide oxytocin regulates parochial altruism in intergroup conflict among humans. Science, 328, 1408–1411.
Demaree, H. A., Everhart, D. E., Youngstrom, E. A., & Harrison, D. W. (2005). Brain lateralization of emotional processing: Historical roots and future incorporating “dominance”. Behavioral and Cognitive Neuroscience Reviews, 4, 3–20.
Domes, G., Lischke, A., Berger, C., Grossmann, A., Hauenstein, K., Heinrichs, M., & Herpertz, S. C. (2010). Effects of intranasal oxytocin on emotional face processing in women. Psychoneuroendocrinology, 35, 83–93.
Elliot, A. J., & Thrash, T. M. (2004). The intergenerational transmission of fear of failure. Personality and Social Psychology Bulletin, 30, 957–971.
Ellis, B. J., Boyce, W. T., Belsky, J., Bakermans-Kranenburg, M. J., & Van IJzendoorn, M. H. (2011). Differential susceptibility to the environment: An evolutionary-neurodevelopmental theory. Development and Psychopathology, 23, 7–28.
Euser, E. M., Van IJzendoorn, M. H., Prinzie, P., & Bakermans-Kranenburg, M. J. (2010). Prevalence of child maltreatment in the Netherlands. Child Maltreatment, 15, 5–17.
Feldman, R., Gordon, I., & Zagoory-Sharon, O. (2010). The cross-generation transmission of oxytocin in humans. Hormones and Behavior, 58, 669–676.
Fox, N. A., Henderson, H. A., Rubin, K. H., Calkins, S. D., & Schmidt, L. A. (2001). Continuity and discontinuity of behavioral inhibition and exuberance: Psychophysiological and behavioral influences across the first four years of life. Child Development, 72, 1–21.
Goldstein, M., & Heaven, P. C. L. (2000). Perceptions of the family, delinquency, and emotional adjustment among youth. Personality and Individual Differences, 29, 1169–1178.
Gray, J. R., Braver, T. S., & Raichle, M. E. (2002). Integration of emotion and cognition in the lateral prefrontal cortex. PNAS, 99, 4115–4120.
Hagemann, D., Hewig, J., Seifert, J., Naumann, E., & Bartussek, D. (2005). The latent state-trait structure of resting EEG asymmetry: Replication and extension. Psychophysiology, 42, 740–752.
Harmon-Jones, E., & Allen, J. J. B. (1997). Behavioral activation sensitivity and resting frontal EEG asymmetry: Covariation of putative indicators related to risk for mood disorders. Journal of Abnormal Psychology, 106, 159–163.
Harmon-Jones, E., Gable, P. A., & Peterson, C. K. (2010). The role of asymmetric frontal cortical activity in emotion-related phenomena: A review and update. Biological Psychology, 84, 451–462.
Heim, C., Young, L. J., Newport, D. J., Mletzko, T., Miller, A. H., & Nemeroff, C. B. (2008). Lower CSF oxytocin concentrations in women with a history of childhood abuse. Molecular Psychiatry, 14, 954–958.
Heinrichs, M., Baumgartner, T., Kirschbaum, C., & Ehlert, U. (2003). Social support and oxytocin interact to suppress cortisol and subjective responses to psychosocial stress. Biological Psychiatry, 54, 1389–1398.
Heinrichs, M., von Dawans, B., & Domes, G. (2009). Oxytocin, vasopressin, and human social behavior. Frontiers in Neuroendocrinology, 30, 548–557.
Herrington, J. D., Mohanty, A., Koven, N. S., Fisher, J. E., Stewart, J. L., Banich, M. T., … Heller, W. (2005). Emotion-modulated performance and activity in left dorsolateral prefrontal cortex. Emotion, 5, 200–207.
Hoffman, M. L., & Saltzstein, H. D. (1967). Parent discipline and the child’s moral development. Journal of Personality and Social Psychology, 5, 45–57.
Huffmeijer, R., Alink, L. R. A., Tops, M., Grewen, K. M., Light, K. C., Bakermans-Kranenburg, M. J., & Van IJzendoorn, M. H. (2011). The impact of oxytocin administration and maternal love withdrawal on event-related potential (ERP) responses to emotional expressions with performance feedback. Manuscript submitted for publication.
Huffmeijer, R., Tops, M., Alink, L. R. A., Bakermans-Kranenburg, M. J., & Van IJzendoorn, M. H. (2011b). Love withdrawal is related to heightened processing of faces with emotional expressions and incongruent emotional feedback: Evidence from ERPs. Biological Psychology, 86, 307–313.
Kanat-Maymon, M., & Assor, A. (2010). Perceived maternal control and responsiveness to distress as predictors of young adults’ empathic responses. Personality and Social Psychology Bulletin, 36, 33–46.
Kemp, A. H., & Guastella, A. J. (2011). The role of oxytocin in human affect: A novel hypothesis. Current Directions in Psychological Science, 20, 222–231.
Kobayashi, S., Lauwereyns, J., Koizumi, M., Sakagami, M., & Hikosaka, O. (2002). Influence of reward expectation on visuospatial processing in macaque lateral prefrontal cortex. Journal of Neurophysiology, 87, 1488–1498.
Koenig, A. L., Cicchetti, D., & Rogosh, F. A. (2004). Moral development: The association between maltreatment and young children’s prosocial behaviors and moral transgressions. Social Development, 13, 87–106.
Laufs, H., Krakow, K., Sterzer, P., Eger, E., Beyerle, A., Salek-Haddadi, A., & Kleinschmidt, A. (2003). Electroencaphalographic signatures of attentional and cognitive default modes in spontaneous brain activity fluctuations at rest. Proceedings of the National Academy of Sciences, 100, 11053–11058.
MacDonald, K., & MacDonald, T. M. (2010). The peptide that binds: A systematic review of oxytocin and its prosocial effects in humans. Harvard Review of Psychiatry, 18, 1–21.
Mayo, J. W., & Tinsley, C. H. (2009). Warm glow and charitable giving: Why the wealthy do not give more to charity? Journal of Economic Psychology, 30, 490–499.
McGowan, P. O., Sasaki, A., D’Alessio, A. C., Dymov, S., Labonté, B., Szyf, M., … Meany, M. J. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nature Neuroscience, 12, 342–348.
Meinlschmidt, G., & Heim, C. (2007). Sensitivity to intranasal oxytocin in adult men with early parental separation. Biological Psychiatry, 61, 1109–1111.
Nowark, M. A., & Sigmund, D. (2005). Evolution of indirect reciprocity. Nature, 437, 1291–1298.
Patrick, R. B., & Gibbs, J. C. (2007). Parental expression of disappointment: Should it be a factor in Hoffman’s model of parental discipline? Journal of Genetic Psychology, 168, 131–145.
Perry, A., Bentin, S., Bartal, I. B. A., Lamm, C., & Decety, J. (2010). “Feeling” the pain of those who are different from us: Modulation of EEG in the mu/alpha range. Cognitive, Affective, & Behavioral Neuroscience, 10, 493–504.
Pizzagalli, D. A., Sherwood, R. J., Henriques, J. B., & Davidson, R. J. (2005). Frontal brain asymmetry and reward responsiveness: A source localization study. Psychological Science, 16, 805–813.
Renk, K., McKinney, C., Klein, J., & Oliveros, A. (2006). Childhood discipline, perceptions of parents, and current functioning in female college students. Journal of Adolescence, 29, 73–88.
Schludermann, E. H., & Schludermann, S. M. (1988). Children’s Report of Parent Behavior (CRPBI-108, CRPBI-30) for older children and adolescents (Tech. Rep.). Winnipeg, Manitoba, Canada: University of Manitoba, Department of Psychology.
Soenens, B., Vansteenkiste, M., Luyten, P., Duriez, B., & Goossens, L. (2005). Maladaptive perfectionistic self-representations: The mediational link between psychological control and adjustment. Personality and Individual Differences, 38, 487–498.
Suske, D. H., & Gallagher, L. (2009). Dopaminergic-neuropeptide interactions in the social brain. Trends in Cognitive Sciences, 13, 27–35.
Thibodeau, R., Jorgensen, R. S., & Kim, S. (2006). Depression, anxiety, and resting frontal EEG asymmetry: A meta-analytic review. Journal of Abnormal Psychology, 115, 715–729.
Tomarken, A. J., Davidson, R. J., & Henriques, J. B. (1990). Resting frontal brain asymmetry predicts affective responses to films. Journal of Personality and Social Psychology, 59, 791–801.
Van IJzendoorn, M. H., Bakermans-Kranenburg, M. J., & Ebstein, R. P. (2011). Methylation matters in child development: Toward developmental behavioural epigenetics. Child Development Perspectives 5, 305–310.
Van IJzendoorn, M. H., Bakermans-Kranenburg, M. J., Pannebakker, F., & Out, D. (2010a). In defence of situational morality: Genetic, dispositional and situational determinants of children’s donating to charity. Journal of Moral Education, 39, 1–20.
Van IJzendoorn, M. H., Caspers, K., Bakermans-Kranenburg, M. J., Beach, S. R. H., & Philibert, R. (2010b). Methylation matters: Interaction between methylation density and serotonin transporter genotype predicts unresolved loss or trauma. Biological Psychiatry, 68, 405–407.
Van IJzendoorn, M. H., Huffmeijer, R., Alink, L. R. A., Tops, M., & Bakermans-Kranenburg, M. J. (2011). The impact of oxytocin administration on donating behavior: Moderation by parental love withdrawal. Frontiers in Developmental Psychology. doi:10.3389/fpsyg.2011.00258
Verhaert, G. A., & Van den Poel, D. (2011). Empathy as added value in predicting donation behavior. Journal of Business Research, 64, 1288–1295. doi:10.1016/j.jbusres.2010.12.024
Wallis, J. D., & Miller, E. K. (2003). Neuronal activity in primate dorsolateral and orbital prefrontal cortex during performance of a reward preference task. European Journal of Neuroscience, 18, 2069–2081.
Zak, P. J., Kurzban, R., & Matzner, W. T. (2005). Oxytocin is associated with human trustworthiness. Hormones and Behavior, 48, 522–527.
Zak, P. J., Stanton, A. A., & Ahmadi, S. (2007). Oxytocin increases generosity in humans. PLoS One, 11, e1128.
Author note
M.T. was supported by Veni Grant 451-07-013 from the Netherlands Organization for Scientific Research (NWO). M.J.B.-K. and M.H.v.IJ. were supported by research awards from the NWO (M.H.v.IJ.: NWO SPINOZA prize; M.J.B.K.: VIDI Grant 452-04-306, VICI Grant 453-09-003).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Huffmeijer, R., Alink, L.R.A., Tops, M. et al. Asymmetric frontal brain activity and parental rejection predict altruistic behavior: Moderation of oxytocin effects. Cogn Affect Behav Neurosci 12, 382–392 (2012). https://doi.org/10.3758/s13415-011-0082-6
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-011-0082-6