


Abstract
The repeated administration of methamphetamine (METH) can result in long-lasting decreases in dopamine (DA) levels, tyrosine hydroxylase activity and DA uptake sites in the striatum. However, whether these changes lead to functional alterations in the dynamics of DA release and uptake has not been extensively examined. The present study usedin vivo electrochemistry and microdialysis to examine potassium- and amphetamine-evoked release of DA in the striatum and nucleus accumbens (NAc) of METH-treated rats. Male Fischer-344 rats were administered METH (5 mg/kg s.c.) or saline four times in 1 day, at 2-hr intervals. One week later the animals were anesthetized with urethane and prepared for in vivo electrochemical recordings. The METH treatment resulted in dramatic decreases in potassium-evoked release of DA and in the rate of DA clearance in the striatum, whereas the NAc was not significantly affected. In vivo microdialysis studies demonstrated significant decreases in basal DA levels and in potassium- and amphetamine-evoked overflow of DA in the striatum of METH-treated animals. Basal and evoked DA levels in the NAc were not altered. Post-mortem levels of tissue DA were decreased by 41 to 67% in the striatum and 25 to 31% in the NAc. These results indicate that the striatum is more sensitive than the NAc to the neurotoxic effects of METH, both in measures of functional dynamics of DA signaling and in tissue levels of DA. It remains to be determined whether these functional changes in DA release and uptake are permanent or tend to recover over time.
METH is a potent psychomotor stimulant that has high potential for abuse in humans (Miller and Hughes, 1994). METH is also a neurotoxin. A single large dose or multiple smaller doses of METH can produce long-lasting decreases in DA content, DA uptake and TH activity in DA terminal fields (Gibb et al., 1994: Seiden and Ricaurte, 1987), as well as evidence of nerve terminal degeneration and gliosis in the striatum (Bowyeret al., 1994; Lorez, 1981; Pu and Vorhees, 1995; Ricaurteet al., 1982). The time course of changes in DA systems is relatively long lasting, and in monkeys (Finnegan et al., 1982; Seiden et al., 1975/76) and rats (Bittner et al., 1981) there are still decreases in striatal DA 6 months after METH administration. Although it is not known whether METH has neurotoxic effects on DA systems in humans, it has been suggested that the large doses of METH taken by abusers may be comparable to neurotoxic doses in animals (Seiden et al., 1988). In a recent study, post-mortem levels of DA, TH and the DA transporter were decreased in striatum from chronic METH users (Wilson et al., 1996), similar to the effects seen in animals. However, levels of dopa decarboxylase and the vesicular monoamine transporter were not significantly decreased. This may indicate that the reductions in striatal DA, TH and DA transporter in the METH abusers were due to METH-induced down-regulation and may not represent permanent degeneration. In any case, the abuse potential of METH and its neurotoxic effects make METH an important drug from the standpoint that chronic use by humans may lead to long-term or permanent changes in brain neurochemistry. Thus, elucidation of functional changes in brain neurotransmitter systems after neurotoxic doses of METH in animals is important for understanding the possible consequences of METH abuse.
Most studies examining the lasting effects of METH neurotoxicity have focused on in vitro assays (i.e., TH activity, DA content and [3H]DA uptake). However, Bowyer et al. (1992) found that the accumulation of [3H]DA by striatal slices and the overflow of [3H]DA evoked from this preparation by elevated potassium or METH were not different in slices prepared from METH-treated or control animals. This suggests that, whereas striatal DA levels are reduced, presynaptic mechanisms regulating DA release and uptake are not altered or compensatory changes have occurred to maintain normal DA release and uptake. However, uptake and release of [3H]DA may not necessarily be accurate indicators of endogenous DA release (Herdon et al., 1985). Furthermore, whereas in vitro studies may allow for more precise control over some experimental variables, it is often unclear how in vitro results are related to what occurs in intact animals. Thus, there is a lack of information on thein vivo functional status of DA systems after neurotoxic doses of METH.
The ability of in vivo electrochemistry to monitor DA release and uptake with a high degree of temporal and spatial resolution has been documented by many investigators (for example, Casset al., 1993; Garris and Wightman, 1994; Gerhardt et al., 1995; Gratton and Wise, 1994; Stamford et al., 1991; Suaud-Chagny et al., 1995; Wood et al., 1992). The use of this technique has several advantages over in vitro slices prelabeled with [3H]DA. For example, with in vivo electrochemistry an intact brain with complete neuronal circuitry is studied and endogenous DA overflow is evaluated, rather than that of radiolabeled DA, which may not have an even distribution in releasable pools (Herdon et al., 1985). In addition, the small size of the electrodes and the high speed of recording allow for multiple measurements in a single region and more detailed analyses of release and clearance processes. However, there are some disadvantages with in vivo electrochemistry. For examination of local application of compounds at multiple sites in a single animal, the animal needs to be anesthetized, and the exact concentrations of compounds in the extracellular space after local application cannot be determined as accurately. Nonetheless, in vivo electrochemistry allows for a more detailed analysis of dopaminergic functioning in intact brain.
In the present study, in vivo electrochemistry was used to map out the potassium-evoked release of endogenous DA, and subsequent clearance of DA, in the striatum and NAc of control and METH-treated rats. In addition, potassium and amphetamine-evoked overflow of DA was examined with in vivo microdialysis in the striatum and NAc, to complement and extend the in vivo electrochemistry studies. The results of these experiments suggest that the METH treatment used has a profound effect on functional DA dynamics in the striatum.
Materials and Methods
Animals.
Male Fischer-344 rats (Harlan Sprague Dawley, Indianapolis, IN) weighing 210 to 280 g were used for all experiments. They were housed in groups of two to four under a 12-hr light-dark cycle, with food and water freely available. All procedures for animal use were in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Animal Care and Use Committee at the University of Kentucky.
METH treatment.
Rats were injected s.c. with 5 mg/kg METH-HCl (Sigma Chemical Co., St. Louis, MO) in saline (1 ml/kg), or saline alone (1 ml/kg), four times in 1 day at 2-hr intervals. Some METH-treated rats became lethargic and lost postural control during the treatment period; they were placed on a cold pack for 15 to 20 min to reduce body temperature. Most of these animals regained postural control within 20 min. A few animals were given a second treatment if they lost postural control a second time during the treatment.
In vivo electrochemistry.
One week after treatment with METH or saline, the animals were anesthetized with urethane (1.25–1.5 g/kg i.p.) and placed in a stereotaxic frame. Body temperature was maintained at 37°C by a heating pad coupled to a rectal thermometer. The scalp was reflected, and the skull and dura overlying the frontal cortices were removed bilaterally. A small hole was drilled in the skull over the posterior cortex for the placement of Ag/AgCl reference electrodes. The reference electrodes were secured in place with dental acrylic.
Electrode/micropipette assemblies were constructed by attaching single-barrel micropipettes to each electrode (Friedemann and Gerhardt, 1992). The tips of the micropipettes had outer diameters of 10 to 20 μm, and they were positioned 280 to 300 μm from the tips of the electrodes. The micropipettes were filled with a solution containing 70 mM KCl, 79 mM NaCl and 2.5 mM CaCl2 (pH 7.4). The recording electrodes each contained a single carbon fiber sealed in a glass capillary (fiber diameter, 33 μm; exposed length, 90–150 μm). Before use, all electrodes were coated with Nafion (six to nine times, with drying at 200°C between coats) to increase selectivity of the electrodes for DA over ascorbic acid (Gerhardt et al., 1984). They were then calibrated in vitro at room temperature in 0.1 M phosphate-buffered saline (pH 7.4) containing 250 μM ascorbic acid (Gerhardt et al., 1987). The response of all electrodes used was linear when calibrated with DA up to 20 μM (correlation coefficients ranged from 0.998 to 1.000). The electrodes had a selectivity for DA over ascorbic acid of at least 800:1 and showed a high sensitivity for DA (14–76 nM with a signal-to-noise ratio of 3.0).
High-speed chronoamperometric electrochemical measurements were continuously made at 5 Hz and averaged to 1 Hz using an IVEC-10 system (Medical Systems Corp., Greenvale, NY). The applied oxidation potential was +0.55 V for 100 msec (vs. the Ag/AgCl reference electrodes), and the resting potential was 0.0 V for 100 msec. The oxidation and reduction currents were digitally integrated during the last 80 msec of each 100-msec pulse. Electrode assemblies were positioned in either the medial or lateral dorsal striatum (1.2 mm anterior to bregma, 1.8 or 3.2 mm lateral from midline and 3.5 mm below the surface of the brain). The base-line electrochemical signal was allowed to stabilize (5–10 min), and then the potassium solution was applied by pressure ejection (Picospritzer II; General Valve Corp., Fairfield, NJ) to evoke the release of DA. The volume of fluid injected was monitored by determining the amount of fluid displaced from the micropipette, using a dissection microscope fitted with a reticule eyepiece, and was based on previous calculations that there are ∼250 nl of solution in a 1-mm segment of the micropipette (Friedemann and Gerhardt, 1992). The volume applied was controlled by setting the pressure ejection parameters at a relativity high pressure (20–40 psi), keeping the time of ejection short (100–200 msec) and rapidly repeating application of pressure until the desired volume was applied. After the signal had returned to base- line, the electrode/micropipette assembly was lowered by 0.5 mm. The new base-line was allowed to stabilize and then the potassium solution was applied again. The potassium applications were repeated in 0.5-mm steps to map the striatum and NAc. Both medial and lateral electrode passes were made in each animal on the same side of the brain. The order of the passes was alternated between animals.
In vivo microdialysis.
Animals were treated with either METH or saline as described above and 7 days later were anesthetized with urethane and positioned in a stereotaxic frame. Microdialysis probes (CMA 12 probes, 3-mm-long dialysis membrane, for the striatum; CMA 11 probes, 2-mm-long dialysis membrane, for the NAc; CMA/Microdialysis, Acton, MA) were slowly lowered into either the right or left medial striatum (1.2 mm anterior to bregma and 1.8 mm lateral from the midline, with the tip of the probe 6.5 mm below the surface of the brain) or NAc (1.2 mm anterior to bregma and 2.0 mm lateral from the midline, with the tip of the probe 8.5 mm below the surface of the brain). The probes were perfused continuously, at a rate of 1.2 μl/min, with artificial cerebrospinal fluid containing 145 mM NaCl, 2.7 mM KCl, 1.2 mM CaCl2, 1.0 mM MgCl2, 0.2 mM ascorbic acid and 2.0 mM NaH2PO4, pH 7.4 (Moghaddam and Bunney, 1989). Fractions of dialysate were collected at 20-min intervals. After a 2-hr equilibration period and the collection of three base-line fractions, the perfusate solution was switched to a 100 mM K+ solution (47.7 mM NaCl, 100 mM KCl, 1.2 mM CaCl2, 1.0 mM MgCl2, 0.2 mM ascorbic acid, 2.0 mM NaH2PO4, pH 7.4) for a single 20-min fraction and then switched back to the original perfusate for five additional fractions. d-Amphetamine (100 μM) was then included in the perfusate for one 20-min fraction, followed by five final fractions with normal artificial cerebrospinal fluid. The dialysate samples were either analyzed immediately by HPLC (15 μl injected onto the column) or frozen on dry ice, stored at −80°C and analyzed within 2 days.
Tissue collection and HPLC analysis.
At the end of the experiments the animals were sacrificed, while still anesthetized with urethane, by decapitation. The brains were rapidly removed and chilled in ice-cold saline. A coronal slice of brain, approximately 2-mm thick and containing the striatum and NAc, was made with the aid of a chilled brain mold (Rodent Brain Matrix; ASI Instruments, Warren, MI). The half of the section containing the electrode or dialysis probe tract(s) was immersed in 10% neutral buffered formalin and saved for later verification of electrode and probe placement. The striatum and NAc were dissected from the other half of the slice as a single piece. The NAc was cut away from the striatum, and the striatum was separated into dorsal and ventral portions with a horizontal cut (for the microdialysis experiments, the striatum was retained as a single piece). The substantia nigra was dissected from a similarly made coronal section through the midbrain. The tissue pieces were placed in preweighed vials, weighed and frozen on dry ice. Samples were stored at −80°C until assayed for monoamine content by HPLC.
For determination of monoamine content, the samples were sonicated in 300 μl of cold 0.1 M perchloric acid containing dihydroxybenzylamine as an internal standard. The samples were centrifuged for 5 min at 15,000 × g, and the supernatant was transferred to 0.22-μm Micropure separators (Amicon, Beverly, MA) and centrifuged at 15,000 × g for 1 min. The filtrate was diluted with HPLC mobile phase, and 50 μl was injected onto the HPLC column.
Levels of DA, DOPAC, HVA, serotonin (5-HT) and 5-HIAA were determined by the procedure of Hall et al. (1989). The HPLC system consisted of a Beckman model 118 pump, Beckman model 507 autoinjector and ESA model 5200A Coulochem II electrochemical detector with a model 5011 dual-detector analytical cell (detector 1 set at +350 mV and detector 2 set at −250 mV). A Keystone Hypersil ODS C18column (3-μm particles, 4.6 mm × 100 mm; Keystone Scientific, Bellefonte, PA) was used for separations. Flow rate was 1.6 ml/min and the mobile phase was 0.17 M citrate-acetate buffer, pH 4.1 (containing 50 mg/liter disodium EDTA, 130–140 mg/liter octanesulfonic acid and 7% methanol). Chromatograms were recorded from both detectors using two dual-channel strip chart recorders. Retention times of standards were used to identify peaks, and peak heights were used to calculate recovery of internal standard and amounts of monoamines and metabolites.
Data analysis.
The electrodes used in this study, although relatively insensitive to ascorbic acid due to the Nafion coating, could still detect 5-HT if the levels were high enough. To confirm that the responses detected were due primarily to DA, both the reduction and oxidation currents were recorded and the ratio of the reduction current to the oxidation current was calculated for each K+-induced response. The electrodes used in this study exhibited reduction/oxidation current ratios of 0.4 or greater for DA and ratios of 0.0 to 0.2 for 5-HT (fig. 1). Ascorbic acid, if detected, was not reduced at the potentials used and therefore gave a reduction/oxidation current ratio of 0.0. All of the responses included in the results had reduction/oxidation ratios of at least 0.4, indicating that DA is the predominant compound detected by the electrodes after potassium application. Two parameters of the electrochemical signals were analyzed, i.e., 1) the maximal amplitude of the signals resulting from the application of potassium and 2) the clearance rate of the signals (the slope of the linear declining portion of the signal between the T20 and T60 time points shown in figure 2; the T20 and T60 points represent the time points at which the signal had declined by 20% and 60%, respectively, of the maximal amplitude). The in vivo electrochemistry data were analyzed with a mixed, two-factor analysis of variance. For the microdialysis experiments the data were expressed as the concentration of DA in the dialysate and as percent change from base-line, where base-line was the average value in the three fractions preceding stimulation by excess K+. All probes were calibratedin vitro before use, to determine acceptable probes (recovery of DA at least 25% for striatal probes and 20% for NAc probes). However, values were not corrected for in vitrorecoveries, because uncorrected values may be better correlated with true values (Glick et al., 1994). The microdialysis results were further analyzed with mixed, two-factor analysis of variance. For the tissue HPLC data, the results were expressed as micrograms per gram of wet weight of tissue and were analyzed with two-tailed ttests.

In vitro response of a Nafion-coated carbon fiber electrode to DA, 5-HT and ascorbic acid. The electrode was calibrated with DA before these responses were obtained. Both oxidation (Ox.) and reduction (Red.) current responses are shown for the application of 2 μM DA (final concentration), 2 μM 5-HT and 250 μM ascorbic acid. Each of the three compounds was added to fresh buffer (arrowheads). The reduction/oxidation current ratios (0.66 for DA, 0.17 for 5-HT and 0.0 for ascorbic acid) are typical for this type of electrode. Similar ratios are obtained by in vivoapplication of the compounds (Luthman et al., 1993; W. A. Cass, unpublished observations). All of the electrodes used in this study had reduction/oxidation current ratios of at least 0.4 for DA; for the in vivo responses, only signals with a ratio of 0.4 or greater were included in the data analysis.

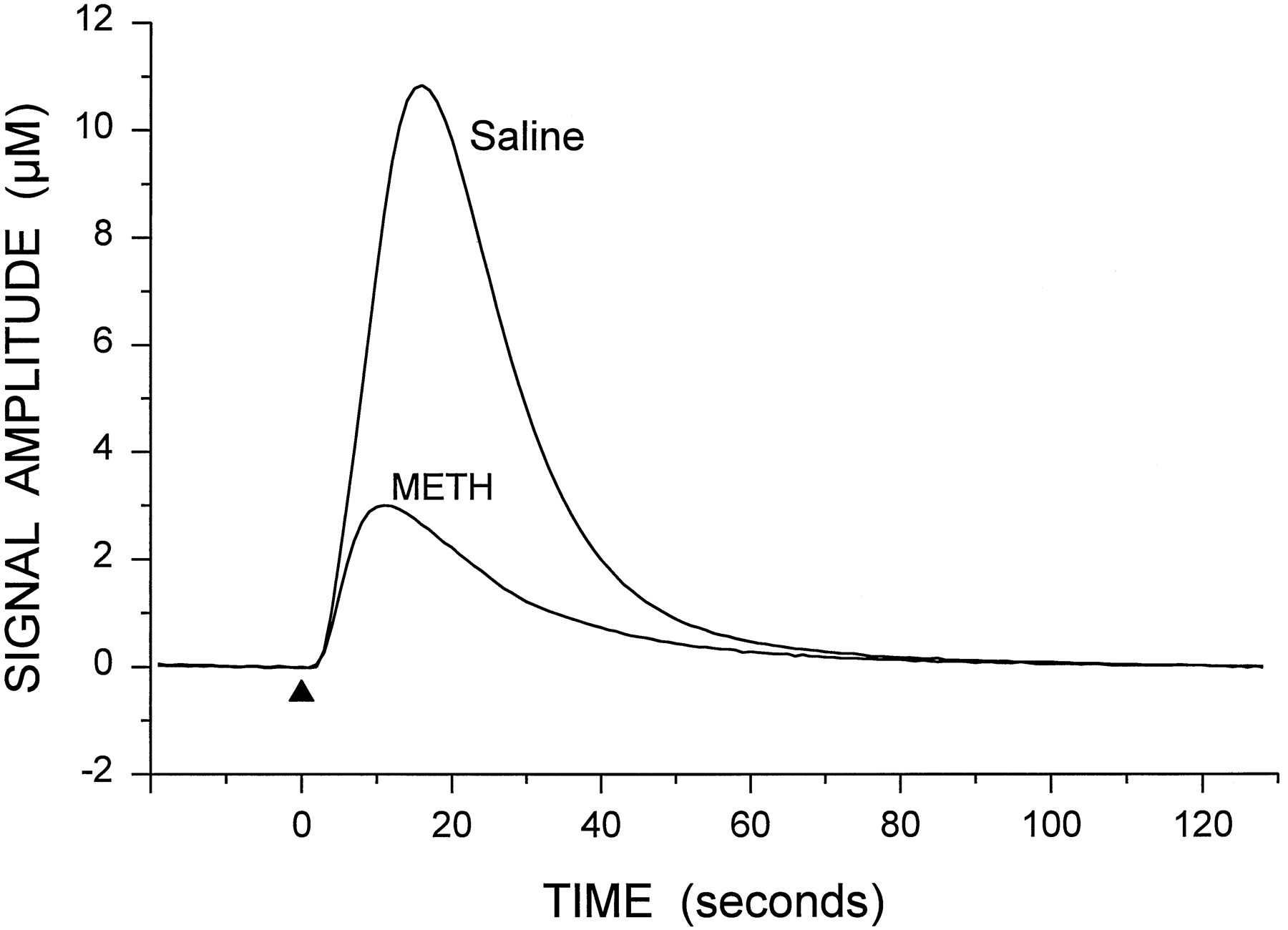
Representative signal showing the potassium-evoked overflow of DA in the striatum of a control rat. Potassium was applied at the arrowhead (250 nl, 70 mM K+). The oxidation (Ox.) and reduction (Red.) current responses (reduction/oxidation ratio of 0.62) indicate that the predominate electroactive species detected is DA. The T20 and T60 time points used in the calculation of clearance rate are shown on the oxidation curve.
Results
In vivo electrochemistry.
The local application of K+ produced DA-like signals in the striatum and NAc of both saline- and METH-treated rats. A representative response, showing the reduction/oxidation current ratio characteristic for DA, from the dorsal striatum of a control rat is shown in figure 2. To determine the optimal amount of K+ to apply to obtain maximal overflow of DA, preliminary experiments were performed in two saline-treated and two METH-treated animals. For these experiments, four or five different volumes of K+ solution (50–450 nl) were applied at the same site in the striatum or NAc. At least 15 min were allowed to pass between applications of K+, to allow the tissue to recover. The order of the volumes applied was varied at each site examined. In all cases 250 to 300 nl produced a maximal response (fig.3). Increasing the volume above 300 nl did not further increase the amplitude of the signals and in some cases decreased the amplitude. This was likely due to dilution of the released DA with the large volume of K+ solution. For the remainder of the experiments, the amount of K+ solution applied at each site was 250 to 300 nl.

Oxidation signal amplitudes after application of four different amounts of K+ solution to a single site in the dorsal striatum of a saline-treated animal and a METH-treated animal. This experiment was repeated at multiple sites in the striatum and NAc of two saline-treated and two METH-treated rats. Although the maximal signal amplitude varied at each site, in all cases the greatest amplitude at any single site was achieved with 250 to 300 nl of K+ solution.
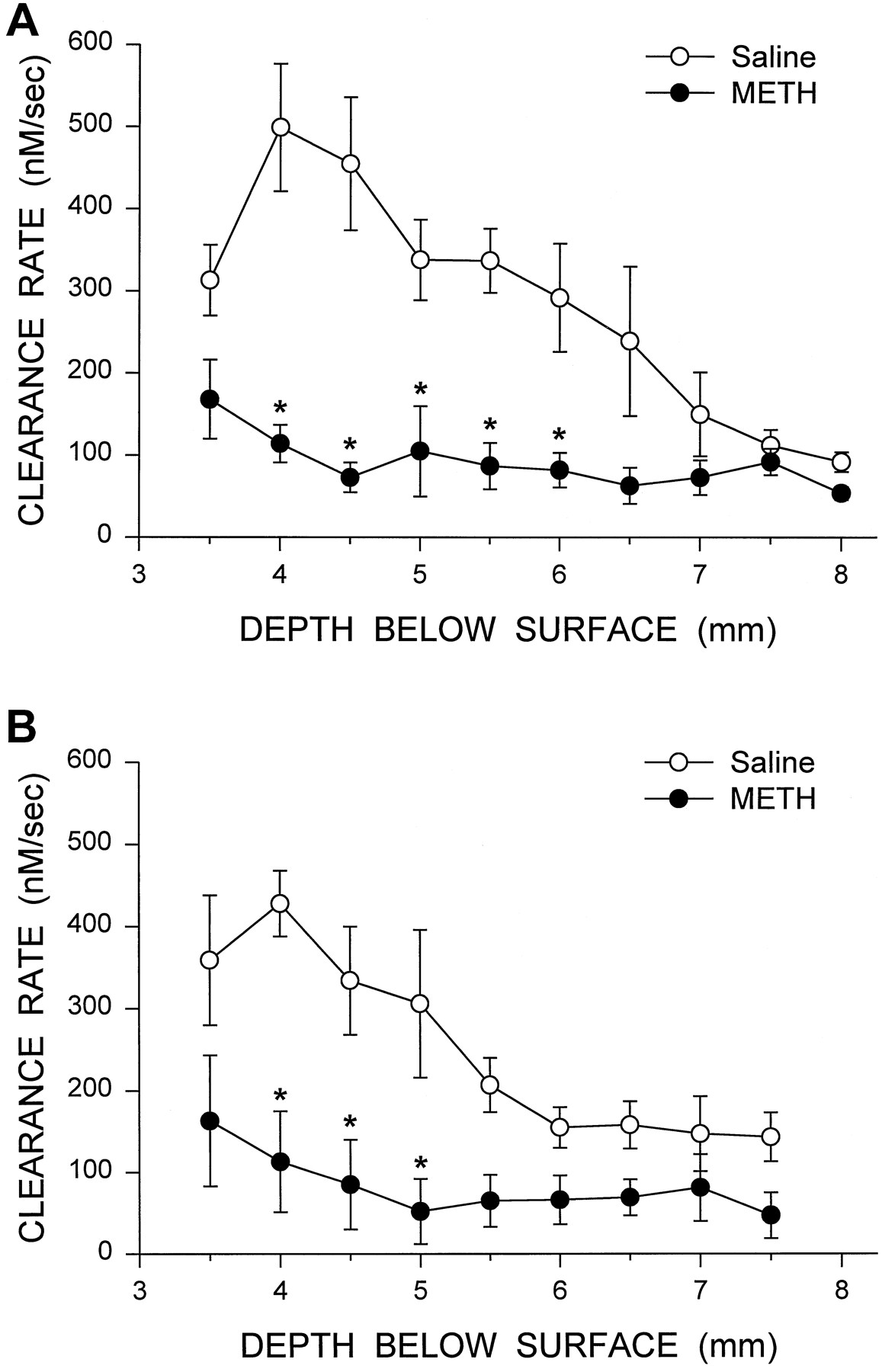
The amplitudes of the potassium-evoked signals were consistently lower in the recordings from the METH-treated animals (fig.4). Signal amplitudes for the medial and lateral passes are shown in figure 5. The histology indicated that for the medial recording pass a depth of 6.5 mm was approximately at the junction of the striatum and NAc, whereas depths of 7.0 and 7.5 mm were contained within the NAc. Recordings at a depth of 8.0 mm were at the ventral edge of, or ventral to, the NAc. For the lateral recording pass, depths of 7.0 and 7.5 mm were within the fundus of the striatum. The METH treatment significantly reduced the amplitude of the signals in both the medial and lateral striatum by >60% (fig. 5). The effects were less pronounced in the NAc. The METH treatment also altered the clearance rate of DA in the medial and lateral striatum (fig.6). Changes in regional values for clearance rate were similar to those for signal amplitude; the dorsal and ventral striatum were affected to a greater degree than the NAc and the fundus of the striatum.

Representative signals for the potassium-evoked overflow of DA from the striatum of saline- and METH-treated animals. Potassium solution (250 nl) was applied at the arrowhead in each case. For clarity, only the oxidation signals are shown.

Summary of potassium-evoked DA signal amplitude throughout the medial (A) and lateral (B) striatum of saline- and METH-treated animals. In the medial recording pass, a depth of 6.5 mm below the surface of the cortex is approximately at the junction of the striatum and NAc, whereas depths of 7.0 and 7.5 mm are within the NAc. The data shown are mean ± S.E.M. values for six saline-treated animals and seven METH-treated animals. The data were analyzed using mixed two-factor analysis of variance, with depth as the within factor. Medial recording F scores: treatmentF = 18.71, P < .002; depthF = 8.23, P < .001; interactionF = 5.71, P < .001. Lateral recordingF scores: treatment F = 12.16, P < .01; depth F = 10.02, P < .001; interaction F = 3.72, P < .001. *P < .05 vs. saline at the same depth (Newman-Keulspost hoc comparisons).

Summary of DA clearance rate after potassium stimulation throughout the medial (A) and lateral (B) striatum of saline- and METH-treated animals. In the medial recording pass, a depth of 6.5 mm below the surface of the cortex is approximately at the junction of the striatum and NAc, whereas depths of 7.0 and 7.5 mm are within the NAc. The data shown are mean ± S.E.M. values for six saline-treated animals and seven METH-treated animals. The data were analyzed using mixed two-factor analysis of variance, with depth as the within factor. Medial recording F scores: treatmentF = 30.13, P < .001; depthF = 7.46, P < .001; interactionF = 5.33, P < .001. Lateral recordingF scores: treatment F = 9.26, P < .05; depth F = 8.13, P < .001; interactionF = 3.61, P < .002. *P < .05vs. saline at the same depth (Newman-Keuls post hoc comparisons).
The tissue levels of monoamines for the animals used in the in vivo electrochemistry experiments are shown in table1. The METH treatment reduced DA levels in the dorsal striatum, ventral striatum and NAc by approximately 41, 67 and 31%, respectively. The DA metabolites DOPAC and HVA were correspondingly reduced in the striatum and NAc. Serotonin also was reduced in the striatum and NAc, and the 5-HT metabolite 5-HIAA was decreased in the dorsal striatum. The METH treatment had no significant effect on the levels of any of these transmitters or metabolites in the substantia nigra (table 1).
Monoamine and metabolite levels 1 week after METH treatment
In vivo microdialysis.
Microdialysis experiments were carried out in the medial striatum and NAc to confirm and extend the results of the electrochemistry experiments. METH treatment 7 days before the microdialysis experiments led to a significant reduction in the amount of DA in the perfusate from the striatum after stimulation by both locally applied potassium and amphetamine. This effect was seen in absolute levels of DA in the perfusate (fig. 7A), as well as in percent change from base-line (data not shown). Basal levels of DA in the perfusate were reduced by 42% and tissue levels of DA were reduced by 49% in the striatum of the METH-treated animals used in the microdialysis experiments (table 2). In contrast, the METH treatment had no significant effect on potassium- and amphetamine-evoked overflow of DA (fig. 7B) or on basal levels of DA (table 2) in the NAc. Tissue levels of DA tended to be lower (by 25%) in the NAc of the METH-treated animals, but the decrease did not reach statistical significance in this group of animals (table 2).

DA levels in dialysate from the medial striatum (A) and NAc (B) of saline- and METH-treated animals. Excess potassium (100 mM) was included in the perfusate for 20 min starting at 0 min, and 100 μM amphetamine was included in the perfusate for 20 min starting at 120 min. The values shown are mean ± S.E.M. from five animals in each group. The data were analyzed using mixed two-factor analysis of variance, with time as the within factor. F scores for the striatum: treatment F = 33.3, P < .001; time F = 65.4, P < .0001; interactionF = 20.4, P < .0001. F scores for the NAc: treatment F = 1.25, P = .29; timeF = 34.5, P < .0001; interactionF = .82, P = .62. *P < .05vs. saline at the same time point (Newman-Keulspost hoc comparisons).
Basal levels of dialysate DA and tissue levels of DA
Discussion
METH is a potent psychomotor stimulant that can also act as a dopaminergic and serotonergic neurotoxin. Many of the earlier reports dealing with the neurotoxic effects of METH on monoaminergic neurons used higher doses of METH than used in the present study, or more extensive treatment schedules (Bittner et al., 1981;Hotchkiss et al., 1979; Ricaurte et al., 1980; Wagner et al., 1979). However, it was subsequently found that a simpler treatment, i.e., four injections spaced 2 hr apart with lower doses of METH, produces a similar depletion of brain monoamines (Bowyer et al., 1992; Eisch et al., 1992; O’Dell et al., 1991; Walsh and Wagner, 1992). Although many studies have documented the DA-depleting effects of METH in the striatum, the effects of neurotoxic doses of METH on functional aspects of presynaptic dopaminergic dynamics have not been extensively examined. The results of the present in vivoelectrochemistry and microdialysis experiments indicate that the METH treatment schedule used in the present study produces extensive reductions in striatal DA release and uptake 1 week after administration of METH.
The decrease in potassium-evoked overflow of DA in the striatum observed with both in vivo electrochemistry and microdialysis indicates a decrease in the depolarization (or calcium)-dependent releasable pool of DA. This is likely due to METH-induced loss of DA terminals. The clearance rate of extracellular DA in the striatum is dependent upon the activity of the DA transporter in the immediate region (Cass et al., 1993). Thus, the decrease in clearance rate in the METH-treated animals indicates loss of DA transporters, as would be expected if DA terminals had degenerated. These results provide functional evidence that METH treatment leads to loss of DA terminals and not just to an extensive depletion of DA stores. Amphetamine-evoked overflow of DA was also examined in the microdialysis experiments. Amphetamine appears to increase extracellular DA in the striatum by a calcium-independent, carrier-mediated, exchange-diffusion mechanism (Kuczenski and Segal, 1994) that has recently been reported to require the presence of the DA transporter (Giros et al., 1996). The decrease in amphetamine-induced displacement of DA in the striatum of the METH-treated rats could be the result of two factors. First, the loss of DA transporters due to terminal degeneration would provide less substrate for amphetamine to act upon. Second, the overall reduction in striatal DA content would provide a smaller pool of DA for displacement by amphetamine. It is likely that both of these factors are involved in the decreased ability of amphetamine to augment extracellular DA levels.
The present results in the striatum are in contrast to those of Bowyeret al. (1992). Those investigators found that the in vitro release of [3H]DA evoked by elevated potassium or METH was not different between striatal slices from saline- and METH-treated animals at 1, 3 or 14 days after METH treatment. However, glutamate-evoked release of [3H]DA was decreased at 1 and 3 days but not 14 days after METH treatment (Bowyer et al., 1992). There are several possibilities for the differing results in their study and the present one. These include measuring endogenous DAvs. [3H]DA and using in vivo vs. in vitro preparations. Another factor is the rat strain used, i.e., Fischer-344 for the present study and Sprague-Dawley for the previous study (Bowyer et al., 1992). It is possible that Fischer-344 rats may be more sensitive to the toxic effects of METH than Sprague-Dawley rats. Although not quantified for this study, there does appear to be behavioral differences during the METH treatment, with the Fischer-344 rats being more excitable and aggressive after the first METH injection (W. A. Cass, unpublished observations).
The present microdialysis results, showing a decrease in basal and evoked overflow of DA in the striatum of METH-treated rats, do not agree with those reported by Robinson et al. (1990). Using microdialysis in freely moving rats, they found decreases in basal levels of DOPAC, HVA and 5-HIAA 1 week after METH treatment. However, basal levels of DA were not decreased, and increases in extracellular DA after systemic administration of amphetamine were not significantly different between METH-treated and control rats. There are many differences between the two studies that could account for these conflicting results. One difference is possible probe placement. In the present study the probe was positioned medially, with its tip at the ventral border of the striatum just above the NAc. This ventromedial placement was chosen because this region was dramatically affected by the METH treatments in the present in vivo electrochemistry and tissue content studies. Although exact coordinates for probe placement are not given in the paper by Robinson et al.(1990), if their probes were positioned more dorsally or laterally this could explain the lack of significant effects on basal and amphetamine-stimulated DA levels, because there are significant variations in the sensitivity to METH in different striatal subregions (Eisch et al., 1992). Another difference is the method of amphetamine stimulation. In the study by Robinson et al.(1990), a submaximal dose of amphetamine (1.5 mg/kg i.p.) was given systemically. For the present experiments, high concentrations of amphetamine (100 μM) or potassium (in both the electrochemistry and microdialysis experiments) were applied locally, to attempt to stimulate maximal release of DA. It is possible that lower concentrations of amphetamine or potassium could have produced less robust differences between the METH- and saline-treated groups. A third difference between the studies is freely moving animals vs.anesthetized animals. The use of anesthesia in the present study may have maximized differences between the METH- and saline-treated groups, possibly by inhibiting compensatory responses. Another variance between the studies that may have affected the results is that they used different rat strains. Overall, there are many differences between the two studies. Additional experiments would be necessary to more fully explain the contrasting results.
Although the results of the METH experiments by Robinson et al. (1990) are not in agreement with the present results, a more recent microdialysis study of 6-hydroxydopamine toxicity from the same laboratory is in agreement with the present findings (Robinson et al., 1994). In rats with partial damage to the nigrostriatal DA system, those authors found a significant decrease in basal extracellular DA concentration in the striatum 4 days after administration of 6-hydroxydopamine. Basal levels returned to normal 3 to 4 weeks after lesioning. Amphetamine-stimulated overflow of DA (1.5 mg/kg i.p.) was also decreased in the partially lesioned animals, at both 4 days and 3 to 4 weeks after lesioning. The extent of the DA depletion was 52 to 54%, similar to the depletion induced by METH in the present study. Thus, these results by Robinson et al.(1994) indicate that partial depletions of DA can lead to significant decreases in basal and evoked release of DA in the striatum of freely moving rats, although over time DA levels may tend to recover.
Most of the literature on METH-induced damage to DA systems has focused on the striatum. However, there are reports that indicate that METH may be toxic to other major terminal regions of mesotelencephalic DA projections, particularly the NAc (Brunswick et al., 1992;O’Dell et al., 1991; Ricaurte et al., 1980;Seiden et al., 1988). In the present study METH reduced the tissue level of DA in the NAc by 25 to 31%. Although the in vivo electrochemistry experiments indicated a trend toward decreased potassium-evoked overflow of DA in the NAc of the METH-treated rats, the decrease was not significant. With the in vivo microdialysis experiments, METH treatment reduced NAc DA levels by a nonsignificant 25% and had no effect on potassium- or amphetamine-evoked overflow of DA. Thus, whereas decreases in tissue DA content were up to 31%, the evoked release of DA was affected minimally or not at all. Taken together, these results indicate that the NAc is less sensitive than the striatum to the neurotoxic effects of METH, both in measures of functional dynamics of DA signaling and in tissue levels of DA.
The results of the electrochemistry experiments in the saline-treated animals demonstrated a dorsal-ventral gradient for potassium-evoked overflow of DA. The signal amplitudes were highest in the dorsal striatum and lowest in the NAc and fundus of the striatum. These differences in amplitude thus correspond closely to tissue content of DA. Changes in the clearance rate of DA in the saline-treated animals paralleled the pattern observed for signal amplitude. The heterogeneity of the distribution of DA transporters in the striatum and NAc (Casset al., 1992; Marshall et al., 1990; Richfield, 1991) is the likely explanation for much of this difference. However, activation of D2 DA receptors can lead to increases in DA uptake by modulating the activity of the DA transporter (Cass and Gerhardt, 1994;Meiergerd et al., 1993; Parsons et al., 1993). This also may play a role in the regional differences in clearance rate. Thus, the activity of the DA transporter would be expected to be higher in the dorsal striatum, where greater extracellular concentrations of DA are achieved after potassium-induced depolarization, and lower in the region of the NAc, where there is less evoked release of DA.
The regional effects of the METH treatment on DA signal amplitude and clearance rate, compared with the saline-treated animals, are interesting. Although there were dorsal-ventral gradients in signal amplitude and clearance rate in the control animals, in the METH-treated animals there were no significant differences at the various recording depths for either signal amplitude or clearance rate. Elevated extracellular levels of DA appear to be required for the neurotoxic effects of METH on DA terminals (O’Dell et al., 1993; Sonsalla et al., 1986; Stephans and Yamamoto, 1994). There may be a threshold level of extracellular DA that needs to be reached for METH to have a neurotoxic effect. The dorsal-ventral gradient in evoked release of DA and in tissue DA levels may explain some of the heterogeneity in the effects of METH. For instance, the lower sensitivity of the NAc to the neurotoxic effects of METH could be due in part to the METH treatment producing a lower cumulative increase in extracellular levels of DA in the NAc, compared with the striatum. Another possibility is that there is a subset of DA terminals, distributed relatively evenly throughout the striatum and NAc, that are less sensitive to the neurotoxic effects of METH. In any case, the METH treatment eliminated the normal gradient for DA release and clearance in the striatum and NAc and thus, in effect, equalized these presynaptic processes across subregions.
In addition to its effects on DA terminals, METH can affect 5-HT systems. The effects on 5-HT are similar to those on DA systems,i.e., decreases in 5-HT content and uptake and tryptophan hydroxylase activity (Gibb et al., 1994). The decreases in 5-HT levels in the striatum and NAc found in the present study are in agreement with this. In addition, similarly to the effects of METH on evoked release of DA in the striatum, the decrease in striatal 5-HT content appears to be reflected as a decrease in evoked overflow of 5-HT when examined by in vivo microdialysis (W. A. Cass, unpublished observations).
The results of the present study indicate that neurotoxic doses of METH produce significant reductions in potassium- and amphetamine-evoked release of DA and in basal levels of extracellular DA. The magnitude of these effects in different brain regions corresponds to the severity of DA depletion. Thus, the effects on DA dynamics are greatest in the striatum, where DA levels are consistently reduced by METH treatment, and minimal in the NAc, which is less sensitive to the DA-depleting effects of METH. It remains to be determined whether these functional changes in DA release and uptake are permanent or tend to recover over time. If these changes are permanent, they could have significant functional consequences over time. For example, METH-induced reductions in striatal DA release could potentially intensify the neurochemical changes in dopaminergic functioning that normally occur during aging (Friedemann and Gerhardt, 1992; Kametani et al., 1995;Marshall and Rosenstein, 1990).
Acknowledgments
I thank Michael Dugan for excellent technical assistance.
Footnotes
-
Send reprint requests to: Wayne A. Cass, Ph.D., Department of Anatomy and Neurobiology, MN 224 Chandler Medical Center, University of Kentucky, Lexington, KY 40536-0084.
-
↵1 This work was supported in part by the University of Kentucky Medical Center Research Fund and by USPHS Grant DA 10115.
- Abbreviations:
- DA
- dopamine
- DOPAC
- 3,4-dihydroxyphenylacetic acid
- 5-HIAA
- 5-hydroxyindoleacetic acid
- HPLC
- high-performance liquid chromatography
- 5-HT
- 5-hydroxytryptamine
- HVA
- homovanillic acid
- METH
- methamphetamine
- NAc
- nucleus accumbens
- TH
- tyrosine hydroxylase
- Received May 2, 1996.
- Accepted September 27, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}