

Abstract
It is well accepted that methylphenidate (MPD) inhibits dopamine (DA) transporter function. In addition to this effect, this study demonstrates that MPD increases vesicular [3H]DA uptake and binding of the vesicular monoamine transporter-2 (VMAT-2) ligand dihydrotetrabenazine (DHTBZ) in a dose- and time-dependent manner in purified striatal vesicles prepared from treated rats. This change did not result from residual MPD introduced by the originalin vivo treatment, because application of MPD in vitro (≤1 μm) was without effect, and higher concentrations decreased vesicular [3H]DA uptake. In addition, MPD treatment increased and decreased VMAT-2 immunoreactivity in striatal vesicle subcellular and plasmalemmal membrane fractions, respectively. The MPD-induced increase in both VMAT-2 immunoreactivity and DHTBZ binding was attenuated by pretreatment in vivo with either the DA D1receptor antagonist SCH23390 or the DA D2 receptor antagonist eticlopride. Coadministration of these antagonists in vivo inhibited completely the MPD-induced increase in DHTBZ binding in the purified vesicular preparation. These observations suggest a role for DA in the MPD-induced redistribution of VMAT-2. The implications of this phenomenon will be discussed.
Methylphenidate (MPD) is one of the most commonly prescribed psychostimulants in the United States. Its primary clinical use is for the treatment of attention deficit hyperactivity disorder (Challman and Lipsky, 2000; Zuddas et al., 2000), which is estimated to affect 3–5% of children in the United States (Pincus et al., 1995). There has been an increase in the illicit use of this stimulant presumably attributable to its pharmacological similarity to other drugs of abuse, such as cocaine. Specifically, MPD inhibits dopamine (DA) transporter function (Ritz et al., 1987; Pan et al., 1994; Izenwasser et al., 1999) and thereby increases extracellular DA levels (Hurd and Ungerstedt, 1989; Butcher et al., 1991).
The vesicular monoamine transporter-2 (VMAT-2) is responsible for the sequestration of cytoplasmic dopamine (Erickson et al., 1992) and is an important regulator of DA neurotransmission; still, no studies to date have investigated the effect of MPD on VMAT-2 function. Hence, the purpose of this study was to determine whether MPD affects VMAT-2 activity. Results reveal that a single administration of MPD rapidly and reversibly increases vesicular [3H]DA uptake and binding of the VMAT-2 ligand [3H]dihydrotetrabenazine (DHTBZ). MPD treatment also increases VMAT-2 protein levels in a striatal vesicle subcellular preparation. These MPD-induced increases in vesicular [3H]DA sequestration, [3H]DHTBZ binding, and VMAT-2 protein levels are mediated by both DA D1 and D2 receptor activation. These phenomena represent an MPD-induced redistribution of vesicles within nerve terminals that may, in turn, alter intraneuronal DA distribution. The implications of this phenomenon are discussed.
MATERIALS AND METHODS
Animals. Male Sprague Dawley rats (280–340 gm; Simonsen Laboratories, Gilroy, CA) were maintained under controlled lighting and temperature conditions, with food and water providedad libitum. Rats were killed by decapitation using a guillotine. Striata (40–50 mg in weight per rat) were dissected and quickly placed in cold 0.32 m sucrose until tissue was processed (for details, see below). All procedures were conducted in accordance with National Institutes of HealthGuidelines for the Care and Use of Laboratory Animalsand approved by the University of Utah Institutional Animal Care and Use Committee.
Drugs and chemicals. (±)MDP hydrochloride was supplied by the National Institute on Drug Abuse (Bethesda, MD). 7,8-[3H]DA (48 Ci/mmol) was purchased from Amersham Life Sciences (Arlington Heights, IL) and α-[2-3H]DHTBZ (20 Ci/mmol) was purchased from American Radiolabeled Chemicals (St. Louis, MO). Tetrabenazine (TBZ) was kindly donated by Drs. Jeffrey Erickson (Louisiana State University Health Sciences Center, New Orleans, LA), Helene Varoqui (Louisiana State University Health Sciences Center) and Erik Floor (University of Kansas, Lawrence, KS). All drugs were administered at 1 ml/kg, as indicated in the figure legends. Doses were calculated as the respective free base, and drugs were dissolved in 0.9% saline.
Preparation of striatal synaptic vesicles. Synaptosomes were prepared from rat striatum as described previously (Fleckenstein et al., 1997). Synaptosomes were then resuspended and homogenized in cold distilled deionized water. Osmolarity was restored by the addition of HEPES and potassium tartrate (final concentration of 25 and 100 mm, respectively, pH 7.5). Samples were centrifuged for 20 min at 20,000 × g (4°C) to remove lysed synaptosomal membranes. MgSO4 (1 mm, final concentration) was added to the supernatant, which was then centrifuged for 45 min at 100,000 ×g (4°C). The resulting vesicular pellet was resuspended in wash buffer (see below) at a concentration of 50 mg/ml (original wet weight of tissue).
Vesicular [3H]DA uptake and [3H]DHTBZ binding. Vesicular [3H]DA uptake was performed by incubating 100 μl (∼2.5 μg of protein) of synaptic vesicle samples at 30°C for 3 min in assay buffer [final concentration (in mm): 25 HEPES, 100 potassium tartrate, 1.7 ascorbic acid, 0.05 EGTA, 0.1 EDTA, and 2 ATP-Mg2+, pH 7.5) in the presence of [3H]DA (30 nmfinal concentration, except in kinetic analyses wherein 0.8–10 μm [3H]DA was used). The reaction was terminated by addition of 1 ml of cold wash buffer (assay buffer containing 2 mmMgSO4 substituted for the ATP-Mg2+, pH 7.5) and rapid filtration through Whatman GF/F filters soaked previously in 0.5% polyethylenimine. Filters were washed three times with cold wash buffer using a Brandel filtering manifold. Radioactivity trapped in filters was counted using a liquid scintillation counter. Nonspecific values were determined by measuring vesicular [3H]DA uptake at 4°C in wash buffer.
Binding of [3H]DHTBZ was performed as described by Teng et al. (1998). Briefly, 200 μl (∼6 μg of protein) of the synaptic vesicle preparation was incubated in wash buffer in the presence of [3H]DHTBZ (2 nm final concentration, except in kinetic analyses wherein 0.25–500 nm DHTBZ was used) for 10 min at 25°C. The reaction was terminated by addition of 1 ml of cold wash buffer and rapid filtration through Whatman GF/F filters soaked in 0.5% polyethylenimine. Filters were washed three times with ice-cold wash buffer. Radioactivity trapped in filters was counted using a liquid scintillation counter. Nonspecific binding was determined by coincubation with 20 μm TBZ. All protein concentrations were determined by a Bio-Rad (Hercules, CA) protein assay.
Preparation of striatal subcellular fractions. Fresh striatal tissue was homogenized in ice-cold 0.32m sucrose and centrifuged (800 ×g for 12 min; 4°C). The resulting supernatant (S1) was then centrifuged (22,000 × g for 20 min; 4°C), and the pellets [P2; whole synaptosomal fraction (plasmalemmal membrane plus vesicular subcellular fractions)] were resuspended in cold distilled deionized water at a concentration of 50 mg/ml (original wet weight of tissue). Resuspended tissue was aliquoted into two test tubes. One aliquot was centrifuged (22,000 × g for 20 min; 4°C) to separate plasmalemmal membranes from the synaptic vesicle-enriched fraction. The resulting supernatant (S3) contained the vesicular subcellular fraction of interest, and the pellets (P3; plasmalemmal membrane fraction) were resuspended in cold distilled deionized water.
Western blot analysis. VMAT-2 antibody was originally kindly donated by Dr. John Haycock (Louisiana State University, New Orleans, LA) and was subsequently purchased from Chemicon (Temecula, CA; AB1767). Binding of VMAT-2 antibody was performed using 60 μl of whole synaptosomal, plasmalemmal membrane, or vesicle subcellular fractions. Samples were added to 20 μl of loading buffer (final concentration, 2.25% SDS, 18% glycerol, 180 mmTris base, pH 6.8, 10% β-mercaptoethanol, and bromophenol blue). Approximately 60 μg of protein of the whole synaptosomal fraction, 40 μg of protein of the plasmalemmal membrane fraction, or 20 μg of protein of the vesicle subcellular fraction was loaded per well in a 10% SDS-polyacrylamide gel. After electrophoresis, samples were transferred to polyvinylidene difluoride hybridization transfer membrane (NEN, Boston, MA). All subsequent incubation steps were performed at room temperature while shaking. Each membrane was first blocked for 2 hr in 100 ml of Tris-buffered saline with Tween (TBST) (250 mm NaCl, 50 mm Tris, pH 7.4, and 0.05% Tween 20) containing 5% nonfat dry milk. Each membrane was then incubated with anti-VMAT-2 antibody (1:4000 dilution) in 13 ml of TBST with 5% milk for 1 hr and then washed five times (two washes for 1 min each and three washes for 5 min each) in 70 ml of TBST with 5% milk. The membranes then were incubated for 1 hr with the goat F(ab′)2 anti-rabbit Ig antibody (Biosource International, Camarillo, CA) at a 1:2000 dilution in TBST with 5% milk. This secondary antibody had been affinity isolated, preabsorbed with human Ig, and conjugated with horseradish peroxidase. The membranes were then washed five times (two times for 1 min each and three times for 5 min each) with 70 ml of TBST and then developed with the Renaissance Western Blot Chemiluminescence Reagent Plus (NEN) according to specifications of the manufacturer. Multiple exposures of blots were obtained to ensure development within the linear range of the film (Kodak Biomax MR; Eastman Kodak, Rochester, NY). Bands on blots were quantified by densitometry measuring net intensity (the sum of the background-subtracted pixel values in the band area) using Kodak 1D image-analysis software.
Data analysis. Statistical analyses among three or more groups were performed using an ANOVA, followed by a Fisher PLSDpost hoc comparison. Analyses between two groups were conducted using a Student's t test. Differences were considered significant if probability of error was <5%.
RESULTS
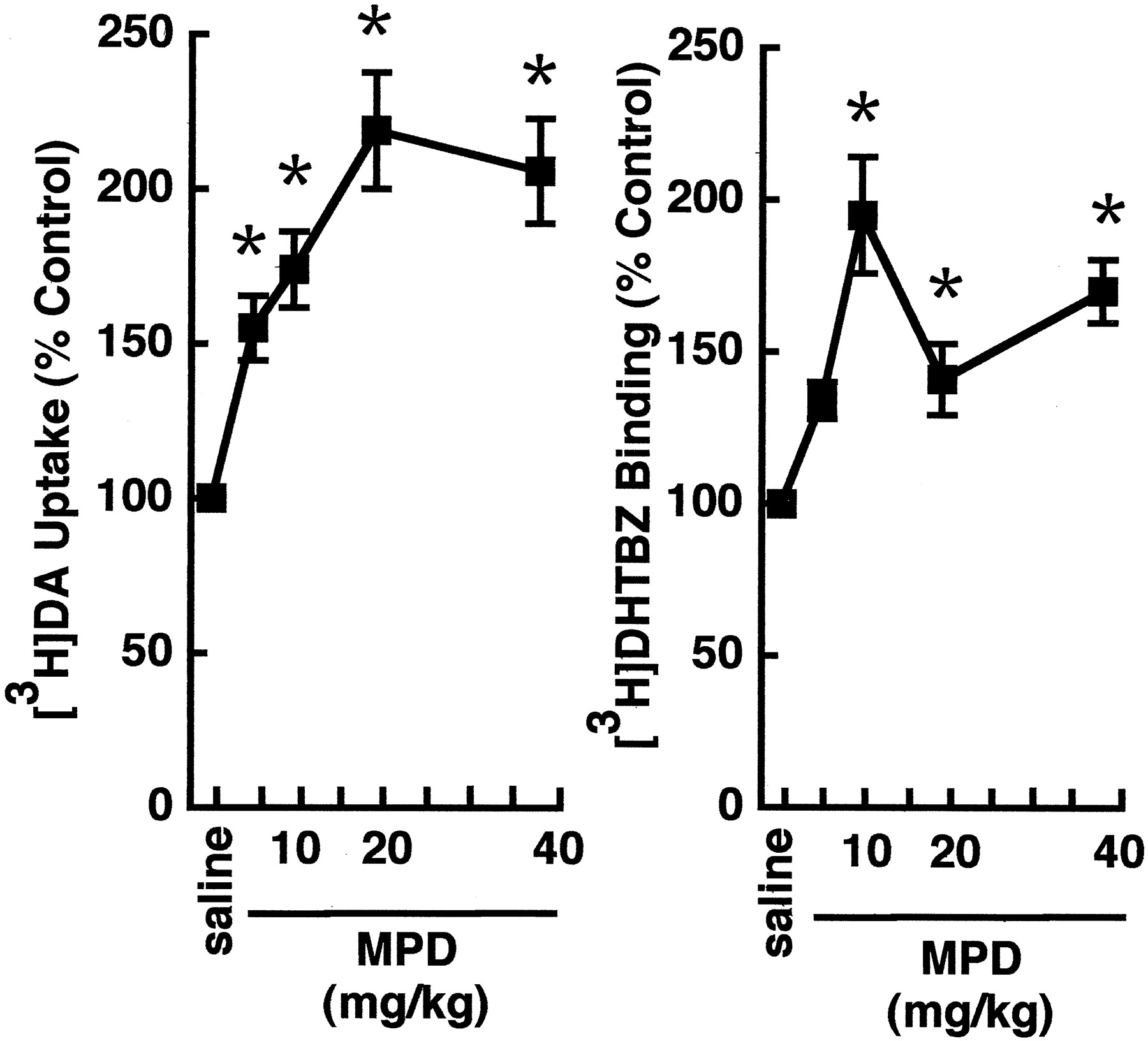
Results presented in Figure1A demonstrate that MPD increases vesicular [3H]DA uptake after a single subcutaneous administration of 5, 10, 20, or 40 mg/kg MPD, as assessed by measuring [3H]DA uptake into purified striatal vesicles prepared from saline- or MPD-treated rats. This increase in vesicular [3H]DA uptake was associated with an increase in binding of the VMAT-2 ligand [3H]DHTBZ (Fig. 1B). The increases in both vesicular [3H]DA uptake and [3H]DHTBZ binding occur rapidly (i.e., within 30 min) and reversibly (i.e., within 12 hr after a 40 mg/kg MPD administration) (Fig. 2). At these doses, MPD administration increased locomotor activity and rearing in the treated animals compared with controls (data not shown).

A single administration of MPD increases vesicular [3H]DA uptake and [3H]DHTBZ binding. Rats received a single administration of MPD (5–40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.) and were killed 1 hr later.Symbols represent the means, and vertical lines represent 1 SEM of determinations in six rats. Data are expressed as a percentage of the mean of control. Mean control values for vesicular [3H]DA uptake and [3H]DHTBZ binding ranged from 81.4 to 167.3 and 1.2 to 2.3 fmol/μg protein, respectively. *p ≤ 0.05, values for MPD-treated rats that are significantly different from saline-treated controls.
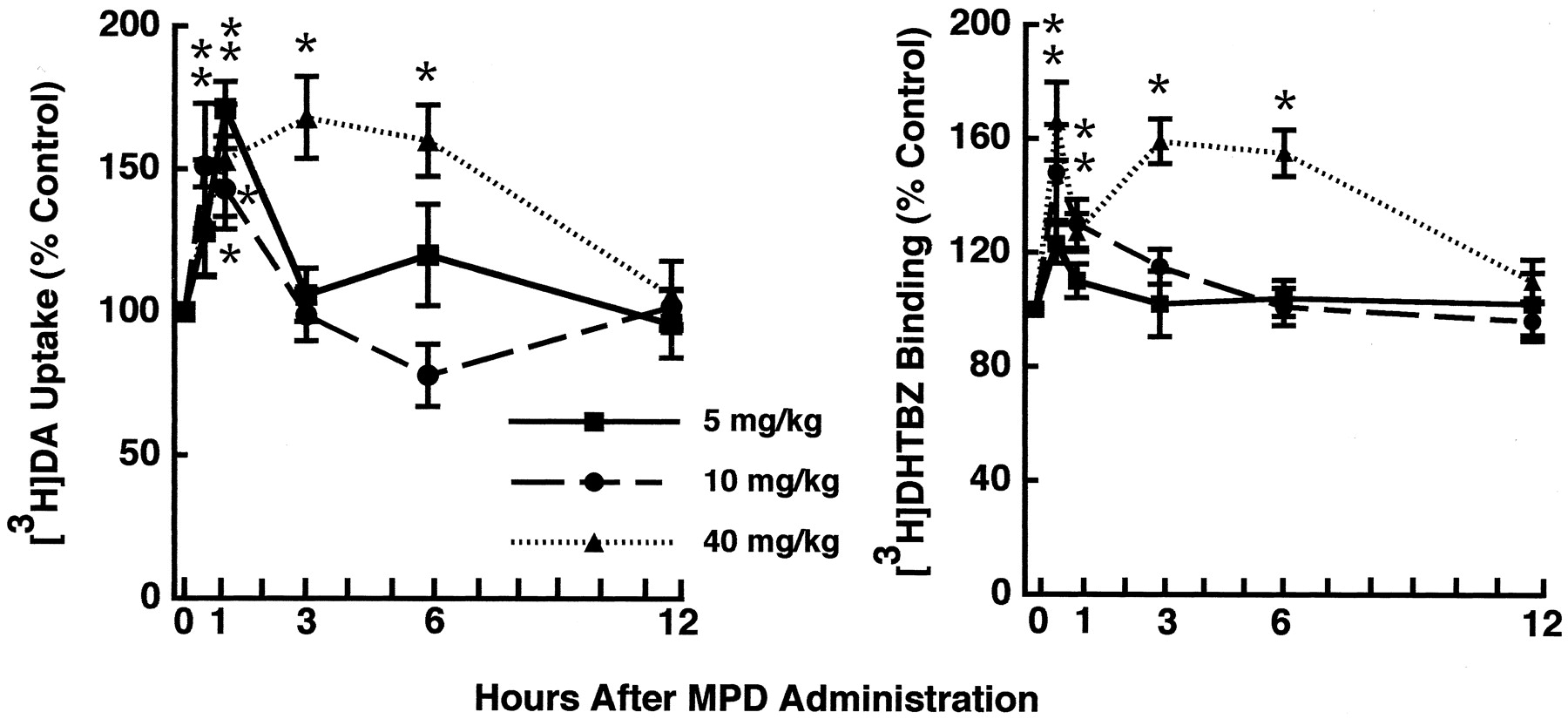
A single administration of MPD rapidly and reversibly increases vesicular [3H]DA uptake and [3H]DHTBZ binding. Rats received a single administration of MPD (5, 10, or 40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.) and were killed 30 min to 12 hr later.Symbols represent the means, and vertical lines represent 1 SEM of determinations in six rats. Data are expressed as a percentage of the mean of control. Mean control values for vesicular [3H]DA uptake and [3H]DHTBZ binding ranged from 135.2 to 226.3 and 4.6 to 7.1 fmol/μg protein, respectively. *p ≤ 0.05, values for MPD-treated rats that are significantly different from saline-treated controls.
The MPD-induced increase in vesicular [3H]DA uptake was associated with an increase in transporter Vmax (for 3 min: 1584 ± 129 and 2350 ± 250 fmol/μg protein for saline- and MPD-treated rats, respectively; p ≤ 0.05) with little change in Km (235 ± 27 and 230 ± 10 nm for saline- and MPD-treated rats, respectively) (Fig. 3). MPD treatment also increased transporterBmax for the VMAT-2 ligand [3H]DHTBZ (18.16 and 28.87 fmol/μg protein for saline- and MPD-treated rats, respectively) with little change in KD (3.02 and 3.25 nm for saline- and MPD-treated rats, respectively). This increase in vesicular [3H]DA uptake did not result from residual MPD introduced by the original in vivo treatment, because direct application of MPD at concentrations of 1 nm to 1 μm was without effect, and higher concentrations of MPD decreased vesicular [3H]DA uptake (i.e., the IC50 for MPD was 19.8 ± 4.0 μm; n = 3).

A single administration of MPD increases theVmax of vesicular [3H]DA uptake. Rats received a single administration of MPD (40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.) and were killed 1 hr later. The Eadie–Hofstee plot depicts data from one of four experiments, with samples in each run in duplicate. The mean Km values were 235 ± 27 and 230 ± 10 nm for saline- and MPD-treated rats, respectively. The mean Vmax values for all four experiments combined were 1584 ± 129 and 2350 ± 250 fmol/μg protein for 3 min for saline- and MPD-treated rats, respectively; these values differed significantly (p ≤ 0.05).
To determine whether the MPD-induced increases in vesicular [3H]DA uptake and [3H]DHTBZ binding were associated with an increase in VMAT-2 protein levels, Western blot studies were conducted in three tissue fractions: vesicular subcellular fraction (i.e., synaptic vesicle enriched), plasmalemmal membrane fraction (i.e., membrane-bound vesicles), and whole synaptosomal fraction (i.e., vesicular subcellular plus plasmalemmal membrane fractions; for detailed description of fractionation, see Materials and Methods). In accordance with data presented in Figures 1 and 2, findings presented in Figure 4Ademonstrate that a single administration of MPD increases VMAT-2 immunoreactivity in the vesicular subcellular fraction. In addition, treatment with MPD decreased VMAT-2 immunoreactivity in the plasmalemmal membrane fraction (Fig. 4B), with no change in the whole synaptosomal fraction (Fig. 4C).

A single administration of MPD redistributes VMAT-2 immunoreactivity. Rats received a single administration of MPD (40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.). All animals were killed 1 hr after the MPD or saline injection. Barsrepresent the mean optic density, and error bars represent the SEM of determinations in six treated rats. Molecular mass standards (in kilodaltons) are shown to the left of the representative Western blot. *p ≤ 0.05, values for MPD-treated rats that are significantly different from saline-treated controls.
To determine whether DA receptor activation contributed to the MPD-induced increases in vesicular transport, [3H]DHTBZ binding, and VMAT-2 protein levels, the DA D1 receptor antagonist SCH23390 or the DA D2 receptor antagonist eticlopride was administered before MPD treatment. Administration of SCH23390 attenuated the MPD-induced increases in vesicular [3H]DA uptake, [3H]DHTBZ binding, and VMAT-2 immunoreactivity in the vesicular subcellular fraction (Fig.5). Moreover, eticlopride pretreatment attenuated the increase in vesicular [3H]DA uptake and completely prevented the MPD-induced increases in [3H]DHTBZ binding and VMAT-2 immunoreactivity in the vesicular subcellular fraction (Fig. 5). Administration of either SCH23390 or eticlopride per se did not affect vesicular [3H]DA uptake or [3H]DHTBZ binding (Figs. 5,6). Coadministration of these antagonists completely inhibited the increase in vesicular DA sequestration and [3H]DHTBZ binding (Fig.7).

A DA D1 receptor antagonist, SCH23390, attenuates the MPD-induced increases in vesicular [3H]DA uptake, [3H]DHTBZ binding, and VMAT-2 immunoreactivity. Rats received a single administration of SCH23390 (SCH; 0.5 mg/kg, i.p.) or saline vehicle (Sal; 1 ml/kg, i.p.) 15 min before a single administration of either MPD (40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.). All animals were killed 1 hr after the last injection.Bars represent the mean vesicular [3H]DA uptake and [3H]DHTBZ binding, and error bars represent the SEM of determinations in six treated rats. Molecular mass standards (in kilodaltons) are shown to the left of the representative Western blot. *p ≤ 0.05, values for MPD-treated rats that are significantly different from saline-treated controls; #p ≤ 0.05, values for SCH23390/MPD-treated animals that are significantly different from MPD-treated animals.

A DA D2 receptor antagonist, eticlopride, attenuates the MPD-induced increases in vesicular [3H]DA uptake, [3H]DHTBZ binding, and VMAT-2 immunoreactivity. Rats received a single administration of eticlopride (Etic; 0.5 mg/kg, i.p.) or saline vehicle (Sal; 1 ml/kg, i.p.) 15 min before a single administration of either MPD (40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.). All animals were killed 1 hr after the last injection.Bars represent the mean vesicular [3H]DA uptake and [3H]DHTBZ binding, and error bars represent the SEM of determinations in six treated rats. Molecular mass standards (in kilodaltons) are shown to the left of the representative Western blot. *p ≤ 0.05, values for MPD-treated rats that are significantly different from saline-treated controls; #p ≤ 0.05, values for eticlopride/MPD-treated animals that are significantly different from MPD-treated animals.

Coadministration of SCH23390 and eticlopride blocks the MPD-induced increases in vesicular [3H]DA uptake and [3H]DHTBZ binding. Rats received a single administration of SCH23390 and eticlopride (SCH & Etic; 0.5 mg/kg, i.p.) or saline vehicle (Sal; 1 ml/kg, i.p.) 15 min before a single administration of either MPD (40 mg/kg, s.c.) or saline vehicle (1 ml/kg, s.c.). All animals were killed 1 hr after the last injection.Bars represent the mean vesicular [3H]DA uptake and [3H]DHTBZ binding, and error bars represent the SEM of determinations in six treated rats. *p ≤ 0.05, values for MPD-treated rats that are significantly different from saline-treated controls.
DISCUSSION
The abuse of MPD has been increasing over the past years; for instance, a 1998 Indiana University survey found that nearly 7% of the students surveyed reported having using MPD illicitly at least once, and 2.5% reported using it monthly or more frequently (United States Department of Justice, Drug Enforcement Administration, 2001). The abuse of MPD can be attributed to its pharmacological similarities to cocaine. In particular, the reinforcing effects of both stimulants are primarily attributable to their ability to inhibit the plasmalemmal DA transporter function. Despite that VMAT-2 has been shown to be an important DA regulator, no studies have been conducted on the effect of MPD on VMAT-2 function. Therefore, the present data demonstrate a novel finding that MPD increases vesicular [3H]DA uptake and [3H]DHTBZ binding rapidly and reversibly, as assessed in purified striatal vesicles prepared from treated rats. These findings are in accordance with previous observations that demonstrated that DA reuptake inhibitors such as cocaine, amfonelic acid, or GBR-12935 increase vesicular [3H]DA uptake and [3H]DHTBZ binding (Brown et al., 2001a). The MPD-induced effects, like the cocaine effects in VMAT-2 function (Brown et al., 2001), are attenuated by pretreatment with eticlopride. Pretreatment with SCH23390 attenuated the MPD-induced increases in vesicular [3H]DA uptake and [3H]DHTBZ binding, but it did not prevent the cocaine-induced increases in VMAT-2 activity (Brown et al., 2001). The coadministration of SCH23390 and eticlopride completely inhibited the MPD-induced increases in VMAT-2 function. Hence, unlike the cocaine phenomenon, both DA D1 and D2 receptor activation contribute to the increase in vesicular [3H]DA uptake and [3H]DHTBZ binding after MPD treatment.
Rapid effects on VMAT-2 have been reported previously after treatment with DA-releasing agents, such as methamphetamine (METH) and methylenedioxymethamphetamine (MDMA). For instance, vesicular [3H]DA uptake and [3H]DHTBZ binding are decreased 1 hr after multiple METH or MDMA administrations (Brown et al., 2000; Hansen et al., 2002). These psychostimulants also acutely decrease plasmalemmal DA transporter function (Fleckenstein et al., 1997; Hansen et al., 2002). These data stand in contrast to our presently reported effect, specifically, that MPD increases (instead of decreases) vesicular DA uptake in our purified vesicular subcellular fraction. Hence, these data demonstrate that vesicular DA uptake can be bi-directionally regulated.
MPD not only increased vesicular [3H]DA uptake and [3H]DHTBZ binding but also levels of the VMAT-2 immunoreactivity in the purified vesicle preparations. One explanation for this may be an increase in the synthesis of the VMAT-2 protein. This possibility is unlikely, because protein synthesis of transporter and receptor proteins typically requires days (Norman et al., 1987; Battaglia et al.,1988; Fleckenstein et al., 1996), a period much longer than the 30 min period at which increases in [3H]DA uptake and [3H]DHTBZ after MPD treatment were observed. A second possibility is an increase generation of functional vesicles. Ongoing studies are addressing this possibility. A third explanation for the MPD-induced increase in vesicular DA uptake, [3H]DHTBZ binding, and VMAT-2 immunoreactivity in the vesicular-enriched subcellular fraction is that these increases are attributable to trafficking of vesicles within the dopaminergic neuron. For instance, MPD treatment may either prevent synaptic vesicle movement from a “cytoplasmic” pool (i.e., purified vesicular subcellular fraction) to the plasma membrane or, inversely, from the plasma membrane to the cytoplasm. Although the present findings cannot definitely discriminate between these possibilities, the data demonstrate that MPD treatment increases and decreases VMAT-2 immunoreactivity in the vesicular subcellular and plasmalemmal membrane fractions, respectively, suggesting that MPD redistributes VMAT-2 protein, and presumably synaptic vesicles, between a subcellular pool and the plasma membrane. In addition, the increase in VMAT-2 immunoreactivity may contribute to the increase inVmax of DA uptake after a single administration of MPD because VMAT-2 protein redistribution, and presumably that of synaptic vesicles, may underlie the increase in the quantity of purified vesicles in our preparation and, thereby, an augmentation in the Vmax of the transporter. However, the possibility that some of theVmax increase might have resulted from an increase in rate (i.e., that the two phenomena are occurring simultaneously) cannot be entirely excluded. It is also relevant to note that, in accordance with results of the [3H]DHTBZ binding studies, DA D1 and D2 receptor activation contribute to the MPD-induced increase in VMAT-2 immunoreactivity in the vesicular subcellular fraction because this increase was prevented by SCH23390 or eticlopride pretreatment (Figs.5, 6). The mechanism(s) underlying this phenomenon remains to be determined.
One possible mechanism whereby MPD treatment causes a redistribution of VMAT-2 protein and presumably synaptic vesicular trafficking may involve presynaptic and/or postsynaptic DA D2receptors. The precise anatomical location of these receptors is unknown, although it is likely that these are ones associated with nigrostriatal regulation. It has been demonstrated that vesicular [3H]DA uptake is increased by administration of the DA D2 receptor agonist quinpirole (Brown et al., 2001b). Interestingly, it has been demonstrated that DA D2 receptors are negatively coupled to cAMP (Stoof and Kebabian, 1981; Vallar and Meldolesi, 1989) and that a decrease in cAMP leads to a decline in protein kinase A (PKA) activation (Beavo et al., 1974). Synaptic vesicles are tethered to cytoskeleton fibers via synapsin, and synapsin is phosphorylated by protein PKA or calmodulin kinase (Turner et al., 1999). Once synapsin becomes phosphorylated, vesicles traffic from the cytoplasm to the plasma membrane (Turner et al., 1999). Consequently, a DA D2 receptor-mediated decrease in PKA activation could cause less synapsin to be phosphorylated and thereby increase the amount of synaptic vesicles tethered to cytoskeletal filaments. This increase in tethered vesicles may underlie the increase in the quantity of purified vesicles in our preparation. This hypothesis does not eliminate the possibility of postsynaptic D2receptor involvement, because DA depletions caused by αMPT treatment can potentially cause decreases in extracellular DA concentrations, and yet αMPT treatment increases vesicular DA uptake (Brown et al., 2001a). Additional investigation regarding the interaction between DA D2 receptors and synaptic vesicles is necessary.
The finding that postsynaptic DA D1receptors contribute to the MPD-induced increase in vesicular [3H]DA uptake, [3H]DHTBZ binding, and VMAT-2 immunoreactivity was unexpected because previous findings by Brown et al. (2001b) demonstrated that administration of a DA D1 receptor agonist, SKF81297, does not increase vesicular [3H]DA uptake or [3H]DHTBZ binding. A possible explanation for the MPD-induced changes may be that DA D1 receptor activation needs to occur concurrently with DA D2 receptor activation for MPD to exert its effects. Future studies are necessary to elucidate any interactions among DA receptors in altering vesicular trafficking.
In conclusion, the data presented demonstrate that a single administration of MPD rapidly and reversibly increases vesicular [3H]DA uptake and [3H]DHTBZ binding by activating both DA D1 and D2 receptors. This phenomenon may result from an MPD-induced redistribution of vesicles within nerve terminals. Pharmacologically altering vesicular trafficking may have important implications beyond explaining differences between the long-term effects of MPD. For instance, autoxidation of cytoplasmic DA has been implicated in the development of Parkinson's disease (Cohen 1990; Fahn and Cohen, 1992; Jenner, 1998; Adams et al., 2001). Accordingly, pharmacological manipulations such as MPD treatment that increase vesicular uptake may be useful in slowing the progression of neurodegeneration by stimulating vesicular removal of DA from potentially unstable cytoplasmic DA pools. Precedence for a neuroprotective sequestering function for VMAT-2 in a neurotoxicity model was suggested previously by the demonstration that VMAT-2 sequesters and thereby protects against the DA neuronal damage caused by the neurotoxin N-methyl-4-phenylpyridinium (Liu et al., 1992; Staal et al., 2000). Interestingly, Falkenburger et al. (2001) demonstrated recently that administration of GBR-12935 [a DA transporter blocker that, like MPD, increases vesicular [3H]DA uptake (Brown et al., 2001a)] abolishes the dendritic release of DA that leads to dendrodendritic autoinhibition. Authors in this study concluded that DA transporter inhibitors may benefit patients in the early stages of Parkinson's disease. Whether a pharmacological enhancement of VMAT-2 function would have clinical relevance remains to be established. However, data presented here suggest that vesicular DA uptake can be regulated and may be a valuable target for treatment of disorders involving disruption of normal DA disposition.
Footnotes
This work was supported by National Institutes of Health Grants DA04222, DA00869, DA11389, and DA13367.
Correspondence should be addressed to Dr. Annette E. Fleckenstein, Department of Pharmacology and Toxicology, 30 South 2000 East, Room 201, University of Utah, Salt Lake City, UT 84112. E-mail:fleckenstein{at}hsc.utah.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}