

Abstract
Background: The sense of agency is an important aspect of motor control. Impaired sense of agency has been linked to several medical conditions, including schizophrenia and functional neurological disorders. A complex brain network subserves the sense of agency, and the right temporoparietal junction is one of its main nodes. In this paper, we tested whether transcranial magnetic stimulation over the right temporoparietal junction elicited behavioural changes in the sense of agency.
Methods: In experiment 1, 15 healthy participants performed a behavioural task during functional MRI, with the goal of localizing the area relevant for the sense of agency in the right temporoparietal junction. In the task, the movement of a cursor (controlled by the participants) was artificially manipulated, and the sense of agency was either diminished (turbulence) or enhanced (magic). In experiment 2, we applied transcranial magnetic stimulation in 20 healthy participants in a sham-controlled, crossover trial with excitatory, inhibitory or sham (vertex) stimulation. We measured the summary agency score, an indicator of the sense of agency (lower values correspond to diminished sense of agency).
Results: Experiment 1 revealed a peak of activation during agency manipulation in the right temporoparietal junction (Montreal Neurological Institute coordinates x, y, z: 68, −26, 34). Experiment 2 showed that inhibition of the right temporoparietal junction significantly reduced the summary agency score in both turbulence (from −14.4 ± 11.4% to −22.5 ± 8.9%), and magic (from −0.7 ± 5.8% to −4.4 ± 4.4%).
Limitations We found no excitatory effects, possibly because of a ceiling effect (because healthy participants have a normal sense of agency) or noneffectiveness of the excitatory protocol.
Conclusion: Our experiments showed that the network subserving the sense of agency was amenable to neuromodulation in healthy participants. This sets the ground for further research in patients with impaired sense of agency.
Clinical trial identification DRKS00012992 (German clinical trials registry).
Introduction
The sense of agency is a fundamental aspect of motor control that allows us to make judgments about whether we performed an action ourselves or not.1 An alteration in the sense of agency has been shown in several disorders, including functional neurological disorders,2 schizophrenia,3 delusions4 or Tourette syndrome;5 patients report a feeling of not being in control of their own actions.6
The sense of agency can be divided into 2 components:7 a low-level, unconscious implicit feeling of being an agent that occurs when actions are simply tagged as self-caused or not; and a higher-level, metacognitive agency in which explicit attributions of agency are made to oneself or another.8
Because of its subjective nature, measurement of both components of the sense of agency represents a methodological challenge.1 Previous research has focused mainly on measurements of implicit sense of agency and has developed well-known paradigms to investigate it, such as the Libet Clock Task9 or the Sensory Attenuation Task.10 The explicit sense of agency has received little attention, and current methods to investigate it typically consist of computer-based games, in which participants first perform a motor task and then explicitly evaluate their sense of control over it.11
Studies into the sense of agency have also focused on identifying the neurobiological network subserving agency.12 Results have shown an involvement of areas devoted to the motor system (ventral premotor cortex, supplementary and pre-supplementary motor areas, cerebellum), as well as to cognition and multimodal information processing, such as the dorsolateral prefrontal cortex, the posterior parietal cortex and the insula.4 In particular, a decrease in implicit sense of agency has been associated with hyperactivity in the right temporoparietal junction (rTPJ).13 It has been hypothesized that the rTPJ is a key node of the so-called comparator model of agency,14 by which intended movements are compared with their actual consequences, and perceptual–motor mismatches are detected to generate the feeling that we are the agent of the performed action.15
In parallel to the advances in neuroimaging, interest has been growing in noninvasive brain stimulation methods because of their potential to modulate neural activity associated with specific cognitive functions.16 Transcranial magnetic stimulation (TMS),16 for instance, has shown successful outcomes in several domains, such as attention,17 visual perception18 or memory.19 In TMS, electromagnetic pulses are used to interfere with normal cortical processing, with consequent behavioural changes. Previous studies have shown modulatory effects of TMS on the sense of agency.20 For instance, several studies have reported that the sense of agency can be disrupted by single-pulse TMS applied to the left21 or right22 inferior parietal cortex, as well as by repetitive TMS over the right inferior parietal cortex at high frequency,23 or over the left superior parietal cortex at low frequency.24 A consensus across the studies has not been achieved yet, and a large variety of target areas and stimulation protocols has been proposed. One pitfall in previous studies was that the target area for TMS was often identified from existing literature, and the behavioural tasks used for target identification were generally different from the ones used to test the effects of stimulation. 22–24 For this reason, the authors could not guarantee that the area targeted by TMS was the same area responsible for the cognitive function being tested. Moreover, the behavioural effects of real TMS were not always distinct from sham stimulation,23 leaving open the possibility of a placebo effect.
To overcome these issues, we performed 2 experiments. First, we used functional MRI (fMRI) to localize the area responsible for the sense of agency in the rTPJ. Second, we applied TMS over this area and studied its modulatory effects on the sense of agency using the same behavioural paradigm used in the first experiment. We adopted a study design that evaluated both the inhibitory and excitatory effects of repetitive TMS, as well as enhanced and decreased sense of agency, in a sham (vertex)–controlled study. The main hypothesis for experiment 1 was that a cluster in the rTPJ activates in response to manipulation of the sense of agency. The main hypothesis for experiment 2 was that inhibitory TMS over the rTPJ would decrease the sense of agency, and excitatory TMS would increase it.
The ultimate translational goal of this study was to develop a TMS-based therapeutic intervention for patients with abnormal sense of agency.
Methods:
Experiment 1: localizer
Participants and ethical approval
Fifteen healthy volunteers (7 men, 8 women; mean age 27.7 ± 4.8 yr) participated in this experiment. All participants were naïve to the hypotheses of the study. Inclusion criteria were as follows: age 16 years or older, and normal or corrected-to-normal vision. Exclusion criteria were as follows: presence of metal clips in the body; history of actual or suspected epilepsy or alcohol or drug abuse; past surgery in the brain; implanted medical devices; and, for female participants, breast-feeding or pregnancy. None of the participants was taking any medication at the time of the study. We did not control specifically for birth control medications. The study was carried out in accordance with the Declaration of Helsinki, and ethical approval was provided by the Ethics Committee of Canton Bern, Switzerland (number 2017–00997). We approached participants 1 week before the experiment, explained the study procedure and obtained written informed consent.
Test setup
The behavioural task used in this experiment was an adaptation of the game proposed by Metcalfe and Greene.11 Participants were shown a pattern of 13 targets and 13 distractors moving downward at a constant speed from the top of a computer screen (Appendix 1, Figure S1, available at jpn.ca/190099-a1). Their goal was to move a cursor on a horizontal bar by clicking on the right or left buttons of an input device with the hand they would normally use to control a computer mouse, and to touch all the targets while avoiding the distractors. After each repetition, participants were asked to rate their performance (judgment of performance; JoP) and their sense of control over the game (judgment of agency; JoA) separately on an 11-point Likert scale from −5 (low JoP/JoA) to +5 (high JoP/JoA). We presented different game phases. In the first, the cursor precisely followed the movement imposed by the input device (baseline). In the second, participants’ feeling of control was artificially reduced by adding random noise to the position of the cursor (turbulence) for 25% of the button clicks, so that the cursor did not always respond to the movement imposed by the participants. In the third, the radius that would count for a “hit” was larger than normal, so that participants were credited with touching a target even if they did not, increasing their performance and the feeling of control (magic).11 To provide visual feedback to the participants, the targets turned green when they were hit by the cursor, and the distractors turned red. As a control condition, participants watched targets and distractors streaming down without performing any button presses (pure visual). As well, participants were told that when the frame of the game was green, they were playing in “easy” mode, and when it was red, they were playing in “hard” mode. However, the actual difficulty did not change, regardless of colour. The duration of each repetition was 15 seconds. We administered a total of 62 repetitions in random order: 11 for baseline, 8 for turbulence and 8 for magic for each of the 2 game modes, as well as 8 for pure visual. Between the repetitions, a central fixation cross appeared for 2 seconds.
Experimental design and fMRI acquisition
Before entering the scanner, participants performed a practice session of the agency task. Once in the scanner, they performed a few more practice rounds, and then the actual session started.
Scans were acquired using a 3 T unit (Magnetom Prisma, Siemens) and a 20-channel head coil. We acquired fMRI data and 1 structural image in a single session. Image volumes consisted of trans-axial slices parallel to the bicommissural plane with a voxel size of 3.0 × 3.0 × 3.0 mm. We acquired fMRI data using an echo-planar imaging (EPI)–blood oxygen level–dependent (BOLD) sequence with the following parameters: repetition time 3000.00 ms, echo time 30.00 ms, flip angle 90°, field of view 192 mm, matrix size 64 × 64, number of acquisitions = 500 functional images. The scan protocol for structural MRI consisted of a T1-weighted, magnetization-prepared rapid acquisition gradient echo (MPRAGE) sequence with the following parameters: repetition time 2330.00 ms, echo time 3.03 ms, flip angle 8°, voxel size 1.0 × 1.0 × 1.0 mm, parallel acquisition techniques factor = 2. During the session, participants were instructed to lie still, keep their hand on an input device with push buttons and use it to perform the agency task.
The duration of the experiment was 30 minutes: 5 minutes to prepare the participant for fMRI, 5 minutes to acquire the structural MRI and 20 minutes for the agency task.
Data analysis and statistics
We implemented the data analysis using statistical parametric mapping (SPM12; www.fil.ion.ucl.ac.uk/spm/).
We implemented image preprocessing as follows. We realigned images to the first image of the first run using iterative rigid-body transformations that minimized the residual sum of the square between the first and subsequent images, and co-registered them to the participants’ anatomic T1-weighted mean images. We then normalized images to the Montreal Neurological Institute (MNI) template (2D spline, voxel size 3 mm), and smoothed them using a Gaussian kernel with a full width at half maximum of 8.0 × 8.0 × 8.0 mm.
For each condition, we modelled a predicted BOLD response to each block with a boxcar function based on the onset and duration of the block convolved with the hemodynamic response function.
In the first-level analysis, we computed the single-participant contrasts for 6 regressors of interest, separated: baseline easy, baseline hard, turbulence easy, turbulence hard, magic easy, magic hard. We incorporated movement corrections (realignment parameters) as covariates (6 nuisance regressors). In the second-level (group) analysis, we implemented a flexible factorial design and random-effects statistics, 25 with all 6 regressors modelled as a single within-participant factor. Finally, to localize the brain areas active during manipulation of agency, we studied the effects of turbulence versus baseline, and magic versus baseline. Other contrasts (e.g., turbulence versus magic, hard versus easy) are not reported here, because they were not relevant to the scope of this study.
We thresholded statistical maps at cluster-forming thresholds of 100 voxels, and we applied a family-wise error correction threshold of 0.050 to peaks and clusters.
Experiment 2: transcranial magnetic stimulation
Participants and ethical approval
Twenty healthy volunteers (10 men, 10 women; mean age 26.3 ± 3.2) participated in this experiment. All participants were naïve to the hypotheses of the study and to the effects of TMS. The recruiting procedures, ethical protocol and inclusion/exclusion criteria were similar to those of experiment 1.
Test setup
The agency task we used in experiment 2 was identical to that of experiment 1, except that participants were seated in front of a computer screen (22 inches; resolution 1680 × 1050 pixels) adapted in height to the level of their eyes.
To control for visual attentional levels, participants also performed a visual detection task.26 For this, they were positioned with their head on a chin and forehead rest 40 cm from the computer screen. While fixating a central cross on the screen, participants had to detect small stimuli that appeared briefly for 40 ms at 24° eccentricity from the fixation cross. The stimuli appeared on the left side, the right side, both sides simultaneously, or not at all. Participants were asked to react to this task as quickly as possible by pressing the mouse button that corresponded to the side on which the stimuli appeared, or on the central scroll when a stimulus appeared on both sides simultaneously. The size of the stimuli was defined as the individual detection threshold of each participant, measured at the beginning of the task.26 We presented a total of 70 repetitions — 20 with stimuli on the right, 20 on the left, 20 on both sides, and 10 with no stimuli — in random order.
Both the agency task and the visual detection task were programmed in Matlab.
Experimental design
This experiment was designed as a single blind, within-subject, vertex-controlled, randomized crossover trial (Appendix 1, Figure S2), with 3 different sessions separated from each other by at least 1 week. After a practice session of the agency task (as described above), the actual task started, followed by the visual detection task with its practice session.26 Then, we applied stimulation (inhibitory, excitatory or vertex stimulation, randomly assigned). Directly after the stimulation, participants performed the agency task and the visual detection task again. At the end of each measurement session, participants were asked about potential adverse effects of TMS, such as nausea, headache or trouble concentrating.27 The duration of each session was 70 minutes: 10 minutes for general assessment, 20 minutes for the first measurement, 20 minutes for the stimulation and 20 minutes for the second measurement.
The second and the third sessions were administered in the same way and took place at the same time of day. The order of the stimulation conditions in the respective sessions was assigned randomly and counterbalanced across participants (i.e., participants who received inhibitory stimulation in the first session received excitatory or vertex in the second, and vertex or excitatory in the third).
Transcranial magnetic stimulation
We administered TMS using a MagPro x100 TMS device (Medtronic Functional Diagnostics) with a MC-B70 butterfly coil. At the beginning of the first session, we measured the motor threshold of each participant using the relative frequency method.28 Then, we selected the target region for the stimulation according to the results of experiment 1, and corresponding to the right supramarginal cortex (MNI coordinates x, y, z: 62, −34, 30). We projected this target point onto the participant’s scalp using the method developed by Steinsträter and colleagues.29 This method has been used in other studies with noninvasive brain stimulation techniques where the individual head model was not available.30,31 We first matched the standard MNI head with the standard 10–20 EEG system by fitting a sphere on both coordinate systems. In this frame, a point described in MNI coordinates corresponds to a point inside the sphere. We projected this point onto the surface of the sphere and reported its coordinates as relative distance from the electroencephalogram (EEG) electrodes (Appendix 1, Figure S3A). We then used a real EEG cap to fit the projected point on the head of the participant (Appendix 1, Fig. S3B). The coil was positioned at a 45° angle toward the contralateral forehead. For vertex stimulation, the coil was positioned over Cz orthogonal to the forehead; this corresponded to stimulation over the interhemispheric fissure (i.e., a brain region not involved in the network of agency).
We based the stimulation patterns for inhibitory and excitatory TMS on continuous and intermittent theta burst stimulation (cTBS and iTBS), introduced by Huang and colleagues32 and modified by Nyffeler and colleagues33 (Appendix 1, Figures S4A and S4B). Briefly, we delivered 2 consecutive applications of cTBS (270 triplets at 30 Hz every 167 ms, for a total of 810 pulses), iTBS (270 triplets at 30 Hz every 167 ms, with an 8.33 s break every 10 triplets, for a total of 810 pulses) or vertex stimulation, with a 15-minute interval between the 2 applications, at an intensity of 80% of the resting motor threshold. Such a protocol has already been proven to extend the duration of the after-effects of TMS by several hours.33
Data analysis and statistics
For analysis of the agency task, we calculated the sense of agency as a summary agency score (SAS), computed from the median JoA and JoP for each game phase separately, and described as:
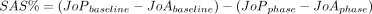
where phase referred to either turbulence or magic.34 This index was negative for decreased agency, and close to zero when full agency was perceived. For the visual detection task, we measured the performance of each participant as the number of detected stimuli in the right, left and bilateral conditions.
We assessed the influence of TMS on the sense of agency using the SAS with repeated-measures analysis of variance (ANOVA) and time (pre, post) × stimulation (cTBS, iTBS, vertex) × game (turbulence, magic) as within-subject factors. We tested the outcomes of the visual detection task with repeated-measures ANOVA and time × stimulation × location (right, left, bilateral). We used Tukey honestly significant difference (HSD) tests for post hoc analyses.35 We calculated effect size as Cohen’s d.36 We analyzed data using STATISTICA 8.0 (StatSoft Inc.).
Results:
Experiment 1: localizer
The results of this experiment are summarized in Table 1 and shown in Figure 1. Three main clusters were significantly activated in the contrast turbulence > baseline: 2 in the right frontal cortex (MNI coordinates x, y, z: 48, 20, 10 and 50, 40, 2), and 1 in the right supramarginal cortex (MNI coordinates x, y, z: 62, −34, 30). No significant cluster was activated in the contrast magic > baseline.

Experiment 1, fMRI results: contrast of turbulence versus baseline. Three main peaks of activation are visible in (A) the right supramarginal gyrus (Montreal Neurological Institute [MNI] coordinates: x, y, z = 62, −34, 30) and (B) the right inferior frontal gyrus (MNI coordinates: x, y, z = 48, 20, 10 and x, y, z = 50, 40, 2).
Experiment 1 — second-level group analysis for the contrast of turbulence versus baseline
Experiment 2: transcranial magnetic stimulation
The results of the repeated-measures ANOVA showed a significant main effect of game: the SAS was significantly lower in the turbulence phase than in the magic phase (F1,19 = 59.55; p < 0.001; d = 3.54). Detailed results are shown in Figure 2.

Experiment 2 results. Summary agency score before and after continuous theta burst stimulation (cTBS), intermittent theta burst stimulation (iTBS) and vertex stimulation, respectively. (A) Summary agency score for the turbulence phase. (B) Summary agency score for the magic phase. Bars represent mean values ± standard error of the mean. *Significant differences according to Tukey honestly significant difference (p < 0.05).
In addition, we found a significant effect of the interaction stimulation × time (F2,38 = 3.60; p = 0.037; d = 0.86). Tukey HSD tests revealed that the SAS significantly decreased after cTBS, from −14.4 ± 11.4% to −22.5 ± 8.9% during turbulence, and from −0.7 ± 5.8% to −4.4 ± 4.4% during magic. We observed no significant changes in the SAS for the iTBS and vertex conditions. Baseline values were not significantly different across stimulation conditions.
We found no significant pre/post differences in the visual detection task in any of the stimulation conditions.
All participants tolerated TMS well: 5 reported mild headache lasting for a few minutes, and 5 reported mild trouble concentrating.
Discussion
Brain areas sensitive to manipulation of agency
The results of experiment 1 showed that when the sense of agency was diminished (turbulence), 3 areas showed hyperactivity: 1 in the rTPJ and 2 in the right inferior frontal gyrus. These results were in line with previous work focusing on the neuronal correlates of sense of agency.13 In particular the rTPJ, as a key node of the comparator model of agency,15 has been found to play a role in detecting mismatches between intended and actual movements.
We recorded no specific brain activation in response to enhanced sense of agency (magic). This suggests that enhanced agency does not have a specific neural signature,37 and the network of agency becomes active only when the sense of agency is decreased.
Effects of TMS on the SAS
Our results showed that inhibitory cTBS decreased the sense of agency in healthy participants (Figure 2). This finding implied that TMS could be successfully used to manipulate the sense of agency when applied over the rTPJ, as identified in our fMRI localizer experiment (MNI coordinates x, y, z: 62, −34, 30). Stimulation over the vertex did not elicit behavioural changes, confirming that our results were not due to a placebo effect, but to a neuromodulatory effect of TMS on the network subserving the sense of agency. A possible alternative explanation of our findings is that TMS induced nonspecific modulatory effects on other cognitive processes in the right posterior parietal cortex, such as visual–spatial attention. 17,38 We addressed this issue by controlling for attentional changes with the visual detection task. Our results showed no difference in attentional levels before and after stimulation, confirming the modulatory effects of TMS specifically on the sense of agency, and not on visual attention.
Our results were in line with those of previous research showing behavioural effects of TMS over the parietal cortex in healthy participants.22–24 In particular, Ritterband-Rosenbaum and colleagues23 showed that 1 second of TMS at 10 Hz over the right inferior parietal cortex induced participants to experience self-controlled movements as externally perturbed. Similarly, MacDonald and colleagues24 found that 15 minutes of TMS at 0.6 Hz over the left superior parietal cortex impaired participants’ awareness of self-generated movements.
In this context, our study brings valuable new elements that increase the evidence of a neuromodulatory effect of TMS over the rTPJ. Previous research has investigated the sense of agency by asking participants whether they thought they were the cause of an action or not.22,23 Instead, we used a Likert scale to measure the sense of agency—a measure that provided not only the direction of the information but also the intensity39 — and this allowed us to measure fine changes in the sense of agency that forced-choice tasks (only direction) may not detect.
Another important behavioural finding of our study was that the sense of control over the game was decreased when the rTPJ was inhibited by TMS. Studies in functional neurologic disorders have shown that patients with hypoactivation in the rTPJ also exhibit decreased sense of agency.40 It is then possible that TMS induced a so-called virtual lesion in the rTPJ, a well-known phenomenon that disrupts task-related neuronal activity in healthy participants,41 simulating the behaviour of patients with pathological hypoactivity. In further research, a direct comparison between patients with reduced rTPJ activity and healthy participants with a virtual lesion in the rTPJ is needed.
Regarding the stimulation protocol, we chose to use 2 consecutive applications of 30 Hz TBS33 instead of 1 application of 50 Hz TBS,32 because previous research has already compared these 2 protocols and found that cTBS at 30 Hz induces stronger inhibitory after-effects in the motor evoked potentials, mainly because of a reduction in inter-participant variability. 42 It has also been shown that the effects of a single application of the 50 Hz protocol are abolished by subsequent voluntary contraction, whereas paired cTBS at 10-minute intervals induced changes in motor evoked potentials that were more resistant.43 For iTBS, we showed that excitatory iTBS produced no behavioural effects, in opposition to our hypothesis. This lack of improvement in the sense of agency might suggest an “enhancement ceiling effect”: that is, the sense of agency of cognitively intact healthy participants cannot be enhanced over their own performance ceiling.44,45 However, an alternative explanation is that the iTBS protocol itself did not elicit any effect. This hypothesis is plausible, because other studies showed that the iTBS protocol had a small effect size and outcomes of large variability,46 often visible only at physiologic levels.32
Limitations
Our study had some limitations. We designed our sample size for experiment 1 based on previous research investigating the sense of agency with neuroimaging techniques,14,47–53 in particular with a study using a similar fMRI task to ours,13 but this sample might not have been representative of the entire population. We did not calculate the effect size of our neuroimaging data because, as stated by Reddan and colleagues,54 standard methods to estimate effect size in neuroimaging studies tend to create bias, which in turn overrate the real effect size. This could have been relevant especially in the magic contrast, where we found no significant cluster of activation. For experiment 2, we calculated the effect size and found that Cohen’s d was 0.86 for our main results on the effects of TMS, which can be considered large.36 We did not use online neuronavigation to place the TMS coil over the target area. This might have affected the results, especially during iTBS, where the coil had to be held in place for more than 4 minutes. However, the results of experiment 1 evidenced a large cluster of activity in the rTPJ of a few square centimetres and, given the 5 cm2 focality of the coil used in our study,55 we decided that a precision in the range of millimetres, potentially achieved with neuronavigation, was not needed. Moreover, the localization method we chose29 has high spatial accuracy56–58 and thus ensured that our target was within the desired cluster.
Another limitation — related to the hypothesis that excitatory iTBS would enhance the sense of agency (which we were unable to show) — was our sample size. The negative findings we reported may have been due to a lack of power. We chose not to increase our sample size, because our study had sufficient power to show a significant effect of inhibitory cTBS. As well, these negative findings could be explained by the fact that the protocol we used has been tested only on cTBS,33 and its iTBS version might not have been effective. Moreover, as stated in the discussion, the inclusion of only healthy participants could have led to a ceiling effect.
In future research, we aim to compare participants with and without decreased sense of agency (such as those with functional neurological disorders) and study the ceiling hypothesis.
Conclusion:
Our results confirmed that one of the key areas for agency processing is the rTPJ and demonstrated that stimulation over that node induced behavioural changes in the sense of agency. Inhibitory cTBS decreased the sense of agency, and we interpreted these findings as modulatory effects of TMS specifically on the sense of agency, because this effect was not seen in the vertex condition. We provided evidence that the rTPJ plays a key role in detecting mismatches between intended action and sensory feedback, in line with the comparator model of agency. Future studies should look at the precise role of the other frontal areas evidenced by experiment 1, which could represent other potential targets for TMSinduced manipulation of agency.
Our study provides solid background for future research aiming at developing new treatment options, based on neuromodulation of the sense of agency by means of TMS, for patients with impairments in agency processing.59–62
Acknowledgements
The authors acknowledge the Swiss National Science Foundation (SNSF), the Fondation Leenaards, the University Hospital Inselspital Bern and the University of Bern for their support and thank all the study participants.
Footnotes
Competing interests: R. Müri declares funding from the Swiss National Foundation, outside of the submitted work. No other competing interests were declared.
Contributors: G. Zito, R. Müri, R. Wiest, M. Holtforth and S. Aybek designed the study. G. Zito and L. Anderegg acquired the data, which G. Zito, K. Apazoglou, R. Müri, R. Wiest, M. Holtforth and S. Aybek analyzed. G. Zito and S. Aybek wrote the article, which all authors reviewed. All authors approved the final version to be published and can certify that no other individuals not listed as authors have made substantial contributions to the paper.
- Received May 21, 2019.
- Revision received October 9, 2019.
- Accepted November 11, 2019.
References
In this issue

{kind=link}
{kind=link}
Article tools






Related Articles
Cited By...
- No citing articles found.