

Abstract
It has been hypothesized that a decrease in the synthesis of new neurons in the adult hippocampus might be linked to major depressive disorder (MDD). This hypothesis arose after it was discovered that antidepressant medications increased the synthesis of new neurons in the brain, and it was noted that the therapeutic effects of antidepressants occurred over a time span that approximates the time taken for the new neurons to become functional. Like antidepressants, exercise also increases the synthesis of new neurons in the adult brain: a 2–3-fold increase in hippocampal neurogenesis has been observed in rats with regular access to a running wheel when they are compared with control animals. We hypothesized, based on the adult-neurogenesis hypothesis of MDD, that exercise should alleviate the symptoms of MDD and that potential mechanisms should exist to explain this therapeutic effect. Accordingly, we evaluated studies that suggest that exercise is an effective treatment for MDD, and we explored potential mechanisms that could link adult neurogenesis, exercise and MDD. We conclude that there is evidence to support the hypothesis that exercise alleviates MDD and that several mechanisms exist that could mediate this effect through adult neurogenesis.
Introduction
Adult neurogenesis refers to the growth of new neurons in the adult brain. Initially, reports of adult neurogenesis were viewed with a great deal of skepticism, because neurogenic activity had been assumed to be limited to the developmental period;1 however, the legitimacy of adult neurogenesis in some brain areas has since gained wide acceptance. It is now clear that some regions of the adult mammalian brain contain populations of active progenitor cells that can give rise to new neurons and glia.2–8 Indeed, adult neurogenesis seems to be a characteristic of all mammalian brains, including human brains.9 Still, fundamental questions about adult neurogenesis have yet to be answered: for example, why does adult neurogenesis occur in only some areas of the mammalian brain, and what purposes do these new neurons serve? Studies of neurogenic activity in the hippocampi of laboratory animals aim to answer some of these questions.
Recently, it has been proposed that a deficit in adult neurogenesis may result, or be somehow involved, in major depressive disorder (MDD) and that the mechanism of action for antidepressant medications may involve promoting neurogenesis. 10–14 The discovery that exercise increases the rate of adult neurogenesis in the hippocampus of mice and rats also suggests a mechanism for the claimed therapeutic effect of exercise on MDD.15,16 In this article, we summarize the process of adult neurogenesis as it is currently understood and briefly describe the evidence that implicates neurogenesis in MDD. We then review studies of the antidepressant effects of exercise on MDD and consider molecular factors that could link adult neurogenesis, exercise and MDD.
Where in the brain does adult neurogenesis occur?
New neurons are not created equally throughout the adult mammalian brain; only certain regions possess populations of active progenitor cells.17–20 Substantial adult neurogenesis occurs in only the dentate gyrus of the hippocampus and the subventricular zone adjacent to the lateral ventricles.3,4,8 It has been estimated that about 9000 new neurons are created in the mature rat dentate gyrus every day.21 The number of new neurons created in the subventricular zones has not yet been estimated, but it is likely to exceed the number created in the dentate gyrus. Although most work regarding adult neurogenesis has been done in rats and mice, current information seems to support both the dentate gyrus and subventricular zone of the lateral ventricles as sites of neurogenesis in both nonhuman primates8,22 and humans.9
The hippocampus is a bilateral limbic structure that plays a role in certain learning and memory processes.23–26 It is composed of 2 substructures that are physiologically distinct (Fig. 1A). One is the cornu Ammonis, whose subfields (CA1, CA2, CA3) contain pyramidal cells that exhibit regional differences in their size, afferent input and efferent projections. The CA1 subfields interlock with the second substructure of the hippocampus, the dentate gyrus. The dentate gyrus is a C-shaped structure that is composed of small, round granule cells. New neurons and glia seem to be created from progenitor cells just below this granule cell layer in what is termed the “subgranular zone” (Fig. 1B).
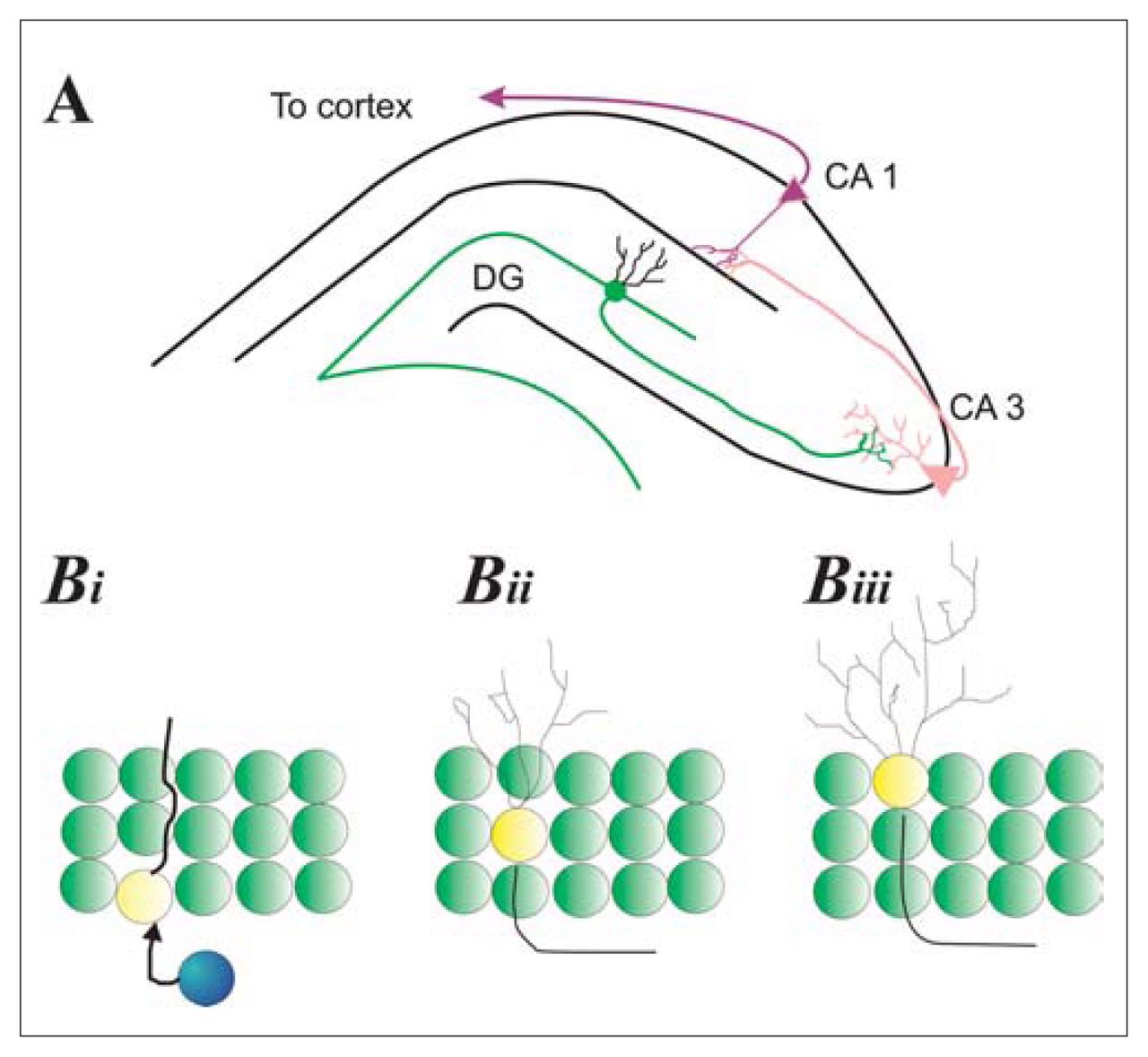
Hippocampal neurogenesis. (A) Schematic of the hippocampus illustrating the relation between granule cells in the dentate gyrus (DG) and pyramidal cells in the cornu Ammonis (CA1, CA3). DG granule cells extend axons from the inner granule zone that project onto dendrites of pyramidal cells in the CA3 layer of the cornu Ammonis (pink). CA3 pyramidal neurons extend axons to CA1 dendrites (black); CA1 cells then extend axons to multiple neural areas, in particular to the cortex. (Bi) Progenitor cells (blue) in the subgranular zone of the DG give way to immature neurons (yellow with extension) that initially extend a proboscis (primary dendrite) through the granular cell layer as well as an axon toward the CA3 region. (Bii) Dendritic complexity increases as neurons mature and migrate into the granule cell layer. (Biii) Some new neurons migrate to the outer granular zone and have dendrites that no longer possess a primary dendrite but, rather, have a more bush-like appearance.
The subventricular region adjacent to the lateral ventricles is the second major source of new neurons in the adult mammalian brain. In this region, there are numerous proliferative precursor cells that develop into neurons or glia.27 These proliferative cells lie just below the layer of ependymal cells, which line the lateral ventricles.28,29
The subgranular and subventricular precursor cells are close to one another in the mammalian brain; the hippocampus forms the medial wall and floor of the lateral ventricle in each hemisphere. However, these 2 sites of adult neurogenesis are characterized by different patterns of development.30,31 Progenitor cells in the subgranular zone divide, and the resulting daughter cells migrate the short distance (20–30 μm) into the granule cell layer. Once in the granule cell layer, the new cells increase the size and complexity of their dendritic tree and send axons to the CA3 region of the pyramidal layer. This process occurs over a period of about 4 weeks, and it results in the formation of new neurons that exhibit morphological and electrophysiological characteristics like those of mature granule cells.31 In contrast, the daughter cells created in the subependymal zones divide, and the resulting daughter cells migrate about 6 mm to the olfactory bulbs (Fig. 1B).28,32 Once in the olfactory bulbs, these new cells become local interneurons. 33
It should be noted that glial cells are also derived from precursor cells in the subependymal and subgranular zones.34,35 Little is known about these new glial cells. It is not known whether they arise from the same precursor cell type as the neurons, or how many new glial cells are born, or what their function may be. Intriguingly, it has also been shown that radial glia themselves may be the progenitor cell in the adult brain.36 Recent demonstrations implicating glial cells in various aspects of neural activity and plasticity suggest that these new glial cells warrant further investigation.37,38
In addition, it has recently been reported that new neurons can, under some conditions, be created in other brain regions, including the neocortex.39–41 However, it remains to be determined whether these become functional neurons.
Environmental influences on adult neurogenesis
There are 3 distinct environmental influences that have been reported to stimulate neurogenesis in adulthood. The first finding was that mice raised in an environment enriched with toys and tunnels had sustained levels of neurogenesis, at least in the dentate gyrus.7 This finding has since been replicated.42,43 The second finding was that learning was capable of stimulating neurogenesis. Rats or mice that were forced to learn a particular task were found to have more new neurons in the dentate gyrus in comparison with rats or mice that did not learn the task.44–46 Interestingly, an investigation into exactly what it was about an enriched environment that increased neurogenesis led to the discovery that voluntary exercise reliably and significantly increases hippocampal neurogenesis.15,16,47,48
Animals that engage in exercise have been reliably shown to undergo a sustained increase in adult neurogenesis in the hippocampus in comparison with control animals that do not engage in exercise15,16,48–50 (Fig. 2). For example, mice given free access to a running wheel for 2–4 months have more than twice the number of new cells in the subgranular zone of the dentate gyrus in comparison with control mice (no access to a running wheel).16 Similar results have also been obtained in rats.47,48 These increases are observed whether animals are housed in small social groups15 or individually.47,48 Increases in adult neurogenesis can be observed as early as 3 days after the start of exercise (unpublished observations) but are more reliably observed after at least a week of exercise.

Exercise increases neurogenesis in adult rats. (Ai–Aiii) Images of 42-day-old cells from a control animal (Ai) and an animal given free access to an exercise wheel (Aii). New neurons are shown in red, whereas mature granule cells are green, and astrocytes are blue. (Aiii) Confocal image of boxed area outlined in red from Ai. For new cells to be considered neurons, they must stain red (bromodeoxyuridine [BrdU], a marker of new cells) and green (Neuronal Nuclei [NeuN], a mature neuronal marker), but not blue (glial fibrillary acidic protein [GFAP], a marker of mature astrocytes). The cell at the centre of the white crosshairs is flipped 90° in both the x and y planes to ensure that the cell is co-labelled for both BrdU and NeuN. (B) Figure shows that exercise (black bars) increases the number of BrdU-positive cells in animals compared with controls (white bars), irrespective of the dose of BrdU administered to the animal (data adapted from Eadie et al, J Comp Neurol 2005;486:39–4747).
MDD and hippocampal pathology
MDD is an affective disorder characterized by depressed mood. This depressed mood can be so severe, enduring and pervasive that it disturbs virtually all daily activities, even such fundamental activities as eating, sleeping and maintaining personal hygiene. About 10% of those diagnosed with MDD commit suicide.51,52 In industrialized Western societies, about 5% of individuals will be diagnosed with MDD at some point in their lives, and the number of people diagnosed with MDD is rising.53,54 Clearly, MDD constitutes a health and social problem of major proportions.55,56
In the last decade, fuelled largely by improvements in brain imaging technology, there has been a concerted effort to identify the neuropathological changes associated with MDD. For example, patients with MDD display a general decrease in cerebral blood flow, as well as reduced glucose metabolism in the amygdala57,58 and the prefrontal cortex.59,60 However, the strongest evidence linking MDD with pathological changes in a specific brain structure involves the hippocampus. 61
Several structural magnetic resonance imaging studies have found hippocampal volume is reduced in depressed patients when compared with age-matched and sex-matched controls.62–68 This reduction in hippocampal volume has recently been found in a postmortem study.69 Although reduced neurogenesis may account for some of the loss of hippocampal volume in patients with MDD, there are likely to be other precipitating factors that may contribute to these structural changes. Stress-induced hormones released from the adrenal gland (e.g., the corticosteroids) can affect the hippocampus by decreasing neurogenesis,70–75 and it has been suggested that these hormones play a role in the decreased hippocampal volume associated with MDD.76–79 Indeed, the stress response is tied to the pathophysiology of MDD; too much stress, or allostatic load, can lead to immune dysfunction, depressed mood and changes in brain structure.80 For example, repeated stress is known to cause atrophy of dendrites in the CA3 region.71 The hippocampus has receptors for adrenal steroids, including the mineralocorticoids and glucocorticoids, 77,81 and it is known to be involved in the negative feedback loop of the hypothalamic–pituitary–adrenal (HPA) axis. This stress response and the effects on hippocampal neurogenesis may also be differentially regulated in males and females.70 Thus, abnormalities in the regulation of the stress response may be related to decreased hippocampal volume in depressed patients,82 which in turn could further affect the stress response.
The adult-neurogenesis hypothesis of MDD
According to the hypothesis that implicates adult neurogenesis in MDD, a decreased rate of neurogenesis contributes to the depressed mood observed in patients with MDD.13,83,84 Three lines of evidence support this hypothesis. First, the hippocampus, which is one of only 2 sites of adult neurogenesis in the mammalian brain, has been found to be smaller in some depressed patients.62–68 Because neural degeneration does not appear to be accelerated in depressed patients, at least in the hippocampus,85 the diminished volume of the hippocampus associated with depression could very well be the product of a reduced rate of adult neurogenesis. Second, adult neurogenesis is increased by treatments for MDD. Antidepressants, such as selective serotonin reuptake inhibitors, have been shown to increase the number of new neurons in the adult hippocampus of laboratory animals, as does electroconvulsive shock treatment.10,86–88 This increase is thought to be the result of the downstream effects of increased serotonergic activation, particularly with regard to brain-derived neurotrophic factor. 89 Third, new neurons normally take 4 or 5 weeks to become functional,31 which is a latency similar to the onset of therapeutic benefit for most programs of antidepressant medication. This suggests that the relief from depressive symptoms may be dependent on the maturation of new hippocampal neurons and their integration into the existing neural network. Although no direct links between depression and adult neurogenesis have yet been demonstrated,90,91 these 3 lines of indirect evidence warrant further evaluation of the relation between adult neurogenesis and MDD.
Effects of exercise on depression
In the 1980s, reports of the beneficial effects of exercise on MDD began to emerge. Most of these published studies were case studies and epidemiological studies, but by 1999 14 studies that had some degree of experimental control had been published. These 14 studies were reviewed by Lawlor and Hopker,92 who concluded that exercise has antidepressant effects that are of the same magnitude as cognitive therapy. However, they tempered their conclusions by pointing out that the 14 studies had a variety of methodological weaknesses. For example, they tended to lack adequate control conditions and blinded outcome ratings, and in most cases the samples were small.92
Subsequent to the review article by Lawlor and Hopker,92 there has been a marked increase in the study of exercise and depression — perhaps in part stimulated by that article. In general, these recent studies have been of higher quality than earlier ones; that is, they have involved larger samples or better controls, or both. Nevertheless, they have confirmed and extended the results of the earlier studies. For example, the recent epidemiological studies by Kritz-Sliverstein et al,93 Motl et al94 and Strawbridge et al95 have shown that young and elderly individuals who engage in programs of exercise display fewer depressive symptoms and are less likely to subsequently develop MDD. In a second series of studies, Leppamaki et al,96 Penninx et al97 and Dimeo et al98 all found that exercise reduces depressive symptoms in adults who have not been diagnosed with a mental disorder.
Most germane to the present article are the recent studies that have assessed the effects of exercise on patients with MDD. Three articles are of particular interest. First, Dimeo et al99 assessed the efficacy of exercise in the treatment of moderate-to-severe MDD. In this study, exercise significantly alleviated the condition. In the second, Mather et al100 assessed the effects of exercise classes as an adjunct to antidepressant medication. The patients who participated in the exercise classes displayed greater improvement than did the control patients, who attended health education talks. Third, Dunn et al101 assessed the effect of the “dose” of exercise on 80 adults with MDD. There were 3 conditions: moderate aerobic exercise, low-intensity aerobic exercise and flexibility exercise. The moderate aerobic exercise group improved significantly more than the other 2 groups.
The results of 2 recent studies are particularly important, because they indicate that the antidepressant effects of exercise endure after the period of exercise. Remarkably, Singh et al102 and Babyak et al103 found that the antidepressant effects of exercise extend past the treatment period with benefits continuing 21 months102 and 6 months103 after the curtailment of the exercise program, respectively.
Another particularly significant recent study of the therapeutic effects of exercise on MDD is the study by Kubesch et al.104 Most previous studies had focused on various measures of depression; however, the various deficits in executive function that appear to be secondary effects of the depression are particularly problematic for many patients with MDD. Kubesch et al assessed executive function by measuring the reaction times of depressed patients on the Stroop task, the Go/No-Go task, the Task-Switching Paradigm and the Flanker task: reaction times on these tests are seen as a measure of executive function. They found that a single 30-minute aerobic endurance exercise period could significantly improve reaction times on these tests in depressed patients. It should be noted, however, that the immediacy of this result would indicate that the benefits of exercise involve more than just increasing neurogenesis.
In summary, recent studies have confirmed earlier (pre-2000) reports that exercise has a significant beneficial effect on depressive symptoms. Accordingly, there is now substantial evidence of the antidepressant effects of exercise. Although there is still a paucity of parametric studies of exercise and MDD, a few recent studies suggest that greater aerobic exercise is associated with greater benefits, that the antidepressant effects of exercise can far outlast the period of exercise, and that the therapeutic benefits in cases of MDD are not solely restricted to depressive symptoms.
Potential mechanisms for exercise-induced neurogenesis
There are several candidate molecules that could play a role in exercise-induced increases in neurogenesis in the adult brain. To find candidate molecules, we screened the literature for molecules (a) whose levels were affected by exercise and (b) that had an effect on the growth of new neurons in the adult hippocampus. The following 4 molecules were selected: β-endorphins, vascular endothelial growth factor (VEGF), brain-derived neurotrophic factor (BDNF) and serotonin (5-HT).
β-Endorphins
An increase in β-endorphins after exercise was first reported over 20 years ago, when β-endorphins emerged as a candidate mechanism to explain why people seemingly become addicted to running.105–108 β-Endorphins are cleaved from the preprohormone pro-opiomelanocortin (POMC), which is a protein found in the pituitary gland and the brain.109 POMC undergoes cleavage to give way to a number of hormones, including the melanocortins and the opiate peptides. β-Endorphins, a class of opiate peptide, share a common N-terminal sequence, function as neuromodulators and are cleaved from the C-terminus of the POMC protein. When β-endorphins bind to their receptors in neural membranes, cyclic adenosine monophosphate (cAMP) levels in the neurons are reduced, and the conductance of voltage-gated Ca2+ channels is decreased.109 Although β-endorphin is not known to cross the blood–brain barrier, levels of plasma β-endorphin-like immunoactivity may indirectly reflect central opioid activity. 110 Importantly, β-endorphins can influence neurogenesis and other hippocampal functions.111,112
Recent studies have shown that an increase in cell proliferation can be produced by the direct infusion of opiates and that opiate receptor antagonists decrease cell proliferation in the dentate gyrus.111,113 Furthermore, when the transcriptional control of enhanced green fluorescent protein (eGFP) is linked to POMC genomic sequences, eGFP is expressed in only new cells in the dentate gyrus.30 Together these results implicate POMC, and possibly β-endorphins, in the genesis, or survival, of new neurons in the dentate gyrus of adult animals.
Vascular endothelial growth factor
VEGF is another molecule that may link exercise and adult neurogenesis.49 VEGF is a protein secreted from blood that acts on endothelial cells to stimulate the formation of blood vessels, and it was first discovered as a vascular permeability factor secreted from cancerous cells.114–116 VEGF family members possess a receptor-binding motif related to that of platelet-derived growth factor.117
VEGF is known to be increased when humans exercise,118 and the infusion of this molecule in the absence of exercise increases neurogenesis in adult rats.119 It is thought that new cells in the dentate gyrus preferentially develop from vascular niches, and it may be the release of VEGF from these blood vessels that leads to an increase in cell numbers in these localized regions.120
Brain-derived neurotrophic factor
BDNF is a member of the neurotrophin family, that is, the same family that includes nerve growth factor and neurotrophins 3 and 4.121 BDNF plays a critical role in the brain throughout development and adulthood by promoting neuronal survival and regeneration.122–125
Exercise leads to increased levels of BDNF.43,126–131 Studies performed in rodents have linked voluntary exercise regimes to increases in BDNF mRNA and BDNF protein. Interestingly, voluntary exercise increases BDNF expression in the central nervous system48,132,133 but not in skeletal muscle,134 indicating that BDNF may play a significant role specifically in the brain. It has recently been found that in “depressed” animals, voluntary exercise can increase both hippocampal cell proliferation and the levels of hippocampal BDNF.135 However, it was concluded that the antidepressant effect of exercise was the result of the enhanced proliferation, because nondepressed animals also exhibited an increase in the levels of BDNF without altering cell proliferation.135 This does not mean, however, that BDNF and exercise do not complement each other in relation to neurogenesis. Indeed, injections of BDNF directly into the hippocampus of rats can enhance proliferation and neurogenesis in the subgranular zone of the dentate gyrus.136 A recent study of mice genetically engineered to express decreased levels of BDNF suggests that BDNF is necessary for the long-term survival of new neurons in the dentate gyrus, because these mice had fewer bromodeoxyuridine (BrdU)-labelled cells 3 weeks after the initial BrdU injection.137 In addition, application of BDNF to cultured dentate gyrus granule cells increases cell survival and differentiation.138 Taken together, these data suggest that exercise might be an effective method for increasing BDNF; BDNF, in turn, can then enhance the survival of newborn cells in the hippocampus.
Serotonin
The link between 5-HT and adult neurogenesis has received a great deal of attention over the past 5 years. The first line of evidence implicating 5-HT in adult neurogenesis was the discovery in rodents that decreases or increases in the brain levels of 5-HT led to decreases and increases in neurogenesis, respectively.86 Currently, specific 5-HT receptor types responsible for regulating both neurogenesis and cell proliferation in the adult brain are beginning to be identified.139 Intriguingly, exercise seems to elevate the levels of tryptophan hydroxylase (the enzyme involved in the rate-limiting step in the synthesis of 5-HT) in the raphe nucleus,140–142 which is an area in the brain stem that is densely populated with serotonergic cells143,144 and sends projections to the hippocampus145,146 that can influence hippocampal activity.147,148 It has also been found that running can increase the levels of tryptophan (which is necessary for the synthesis of 5-HT) in the hippocampus, although this did not increase the levels of 5-HT in the hippocampus.149 Even though there was no specific increase in the levels of 5-HT in the hippocampus, the increased expression of tryptophan might enhance 5-HT production and augment neurogenesis.
Summary
There are several candidate molecules that could play an important role in mediating the effects of exercise on adult neurogenesis. Exercise can increase the levels of β-endorphins, VEGF, BDNF and 5-HT. β-Endorphins might enhance the birth of new neurons in the dentate gyrus, whereas VEGF and BDNF enhance their survival. Exercise also increases the levels of 5-HT in the brain,150–152 which contributes to neurogenesis because 5-HT can increase adult cellular proliferation and neurogenesis in the dentate gyrus.153 Therefore, exercise might have therapeutic effects on MDD, because exercise could increase neurogenesis through the aforementioned putative mechanisms.
Conclusions, limitations and future directions
There is a growing body of evidence that implicates reductions in adult neurogenesis as being associated with MDD. The purpose of this paper was to assess 2 implications of this adult-neurogenesis hypothesis for MDD. First, because exercise has been shown to increase adult neurogenesis in the dentate gyrus, we assessed the evidence for exercise having antidepressant effects. We found substantial evidence that exercise can be an effective treatment for MDD. Second, we reviewed recent studies of adult neurogenesis in laboratory animals to identify molecular mechanisms that might mediate the facilitatory effects of exercise on adult neurogenesis. We identified and discussed 4 molecules that are increased during exercise and have a stimulatory effect on adult neurogenesis: β-endorphins, VEGF, BDNF and 5-HT. These 2 independent lines of evidence together suggest that adult neurogenesis might alleviate the depressive symptoms of MDD.
The major weakness in the empirical support for the adult-neurogenesis hypothesis of MDD is that MDD is a human condition, whereas most of the research on adult neurogenesis has been conducted in nonhuman species. Evidence that exercise increases adult neurogenesis in rodents and decreases depressive symptoms in humans is suggestive but hardly incontrovertible evidence for the hypothesis. In particular, work will need to be done to determine whether the production of new neurons by exercise can play a therapeutic role in alleviating MDD, or whether this phenomenon is merely an unrelated event that is associated with some other factor, such as BDNF, that plays multiple roles in the brain.
Given the inherent difficulties of measuring neurogenesis in human patients, the identification of molecular correlates of adult neurogenesis may prove to be the most beneficial route for future research. However, the evidence thus far provides a hopeful picture for those who have MDD, and further investigations into the role that exercise can play in both structural and functional changes in the brain may help to unveil the nature of MDD in humans and may provide insight into other therapeutic venues for this disorder.
Acknowledgements
This research was supported by Canadian Institutes of Health Research (CIHR), Natural Sciences and Engineering Research Council of Canada (NSERC), Scottish Rite Charitable Foundation of Canada and BC Ministry of Children and Family Development (HELP) awards to Dr. Christie. Mr. Ernst is supported by an NSERC award. Dr. Christie is the BMO Young Investigator at the University of British Columbia Hospital. Dr. Lam has received funding from the CIHR and the Vancouver Hospital Foundation.
Footnotes
Medical subject headings: beta-endorphin; brain-derived neurotrophic factor; depression; exercise; neurogenesis; serotonin; vascular endothelial growth factor.
Competing interests: None declared for Mr. Ernst, Ms. Olson, Dr. Pinel and Dr. Christie. Dr. Lam is on the speaker/advisory boards for AstraZeneca, Biovail, Eli Lilly, GlaxoSmithKline, Janssen, Litebook Company, Inc., Lundbeck, Servier, Shire and Wyeth. He has received research funds from AstraZeneca, Eli Lilly, Glaxo- SmithKline, Janssen, Lundbeck, Merck, Roche, Servier and Wyeth.
Contributors: All the authors contributed to the conception and design of the study. Mr. Ernst and Dr. Christie contributed to the acquisition of the data. All the authors contributed to the analysis of the data and to the writing and critical review of the article. All the authors gave final approval for publication.
- Received August 11, 2005.
- Revision received October 31, 2005.
- Revision received November 11, 2005.
- Accepted November 21, 2005.
References

{kind=link}
{kind=link}
Article tools


