

Abstract
Background: Anxiety disorders are more prevalent in women than in men. Despite this sexual dimorphism, most experimental studies are conducted in male participants, and studies focusing on sex differences are sparse. In addition, the role of hormonal contraceptives and menstrual cycle phase in fear conditioning and extinction processes remain largely unknown.
Methods: We investigated sex differences in context-dependent fear acquisition and extinction (day 1) and their retrieval/expression (day 2). Skin conductance responses (SCRs), fear and unconditioned stimulus expectancy ratings were obtained.
Results: We included 377 individuals (261 women) in our study. Robust sex differences were observed in all dependent measures. Women generally displayed higher subjective ratings but smaller SCRs than men and showed reduced excitatory/inhibitory conditioned stimulus (CS+/CS−) discrimination in all dependent measures. Furthermore, women using hormonal contraceptives showed reduced SCR CS discrimination on day 2 than men and free-cycling women, while menstrual cycle phase had no effect.
Limitations: Possible limitations include the simultaneous testing of up to 4 participants in cubicles, which might have introduced a social component, and not assessing postexperimental contingency awareness.
Conclusion: The response pattern in women shows striking similarity to previously reported sex differences in patients with anxiety. Our results suggest that pronounced deficits in associative discrimination learning and subjective expression of safety information (CS− responses) might underlie higher prevalence and higher symptom rates seen in women with anxiety disorders. The data call for consideration of biological sex and hormonal contraceptive use in future studies and may suggest that targeting inhibitory learning during therapy might aid precision medicine.
Introduction
Anxiety and stress-related disorders are highly prevalent in the general population and are frequently studied in the laboratory by means of fear conditioning and extinction tasks, which serve as models for the acquisition and the behavioural treatment, respectively, of pathological anxiety. During acquisition, a neutral stimulus becomes conditioned (conditioned stimulus [CS]) by repeated pairings with an aversive unconditioned stimulus (US) until the CS has acquired the capacity to elicit a conditioned fear response (CR). During extinction, the CS is presented repeatedly in the absence of the US, which leads to a gradual weakening of the CR that may, however, reoccur at a later time (return of fear [ROF]).1
Anxiety and fear disorders are more than twice as prevalent in women than men,2,3 and affected women also display more severe symptoms and impairment.4,5 Furthermore, data indicate sex differences in the cellular, molecular and structural organization of brain areas involved in emotion and memory (e.g., hippocampus, amygdala, prefrontal cortex6,7). Despite this sexual dimorphism, most experimental fear conditioning studies in rodents have been conducted in male animals. Similarly, the role of sex differences in humans has often been neglected and studies often included only male participants.7,8 When including female participants, studies should control for menstrual cycle phase and the use of hormonal contraceptives because both factors have been shown to have an impact on these brain areas.9 Obviously, this puts a heavy burden on time management and substantially increases the required number of participants. Consequently, only a limited number of studies have investigated the role of sex differences, and all too often conclusions based on studies in male participants are generalized to females.
Biologically determined sex differences in anxiety-related behaviour may arise through different mechanisms.8 They can become hard-wired during early brain development (organizational effects) or arise through phasic variation in sex hormone levels during later phases in life (activational effects). While these mechanisms are not mutually exclusive, support for the latter comes from the observation that sex differences in the prevalence of anxiety do not emerge until puberty10 and that low levels of ovarian hormone levels (e.g., postpartum, premenstrual and perimenopausal) coincide with increased incidences of anxiety symptoms and disorder onset.11,12
Series of rodent studies and studies in naturally cycling females have shown that high levels of estrogens — both occurring naturally during the menstrual cycle and induced pharmacologically — are favourable for extinction memory consolidation, as indicated by enhanced extinction recall. At the same time, low levels have been shown to be disadvantageous for these processes7,8,13–16 and associated with impaired fear inhibition.17 These studies suggest that the consolidation of extinction memory is critically modulated by fluctuating estrogen levels through the menstrual cycle and may be mediated through facilitation of (hippocampal) neuronal plasticity.8 If estrogen in fact modulates the success of experimental fear extinction, this might be of high clinical relevance. Fear extinction is used as a model for exposure-based cognitive behavioural therapy (CBT18), and consequently fluctuations in estrogen levels might affect the outcome of CBT during different menstrual cycle phases.7
The general suppression of estradiol levels by the intake of hormonal contraceptives, which are widely used among young women, might be of similar clinical relevance. It has been suggested that altered fear learning in women taking hormonal contraceptives might be relevant for the treatment of patients with anxiety disorders.19 Previous studies have shown that these contraceptives have little or no effect on the acquisition and extinction of fear but significantly impair extinction recall.20 Furthermore, decreased extinction recall in women and rodents treated with hormonal contraceptives resulting in low estradiol and progesterone levels could be rescued by pharmacological enhancement of estrogen.20 Another study found no differences between men, women using hormonal contraceptives and women in the luteal phase of their menstrual cycles during fear acquisition, whereas during immediate extinction, women taking contraceptives showed significantly stronger excitatory/inhibitory conditioned stimulus (CS+/CS−) differentiation than both of the other groups in regions of the fear network (anterior cingulate cortex, amygdala, thalamus, ventromedial prefrontal cortex) driven by reduced CS− responses.21
Given the markedly different prevalence rates for fear and anxiety disorders between men and women and the wide use of hormonal contraceptives, it is of high clinical relevance whether robust sex differences in fear conditioning and extinction exist and how hormonal contraceptives and menstrual cycle phase modulate these processes. In addition, the role of context dependency in fear learning and extinction processes has not yet been addressed with respect to sex differences. Contexts serve adaptive functions as they help resolve ambiguity and assign situationally dependent meanings. The role of context in fear conditioning, extinction and ROF processes has increasingly gained attention,22–25 and the loss of contextual control over extinction memory has been implicated in psychopathology.25 We thus aimed to investigate differences between men and women and between women who use and do not use hormonal contraceptives in, to our knowledge, the largest sample of healthy participants reported so far. We used a 2-day differential fear conditioning paradigm with context-dependent fear and extinction learning on day 1 and a retrieval/expression session on day 2. This paradigm has been used previously23,26 to study the neural correlates of context-dependent CS discrimination and subsequent extinction retrieval/expression23,26 as well as their pharmacological manipulation.26
Methods
Participants
We recruited healthy individuals (both men and women) who were part of a larger data-collection initiative (Appendix 1, Table S1, available at jpn.ca) to participate in the present study. Additional analyses involved subdividing women into those currently using hormonal contraceptives and free-cycling women and further subdividing free-cycling women based on menstrual cycle phase (follicular v. luteal). We obtained written informed consent from all participants after the procedure had been fully explained. The study was approved by the Ethical Review Board of the General Medical Council of the State of Hamburg (Ethikkommission der Ärztekammer).
Experimental paradigm
The experimental protocol has been used previously.26 In brief, the experiment consisted of a context-dependent fear conditioning and extinction learning task (day 1; 50% reinforcement ratio to make discrimination not too easy) and retrieval/expression of fear and extinction memory (day 2; Fig. 1 and Appendix 1). The paradigm is well suited to address the important role of context dependency in fear learning and extinction processes as fear acquisition often occurs in different contexts (e.g., a traumatic situation) than extinction (e.g., reconfrontation with trauma-associated stimuli outside the traumatic situation in daily life or in therapy). Hence, the paradigm allows us to test whether participants appropriately retrieve fear in the conditioning context and extinction in the extinction context on day 2 or whether the contextualization learned on day 1 might be lost (e.g., fear might generalize to the extinction context, which would be an indication of maladaptive and potentially pathological processing). Prior to the experiment, US intensity was individually adjusted to a level of maximum tolerable pain (range 0.2–85 mA, mean 4.3 ± 0.27 mA). To this end, participants rated the painfulness of the US from 0 (“I feel nothing”) to 10 (“maximally unpleasant”; final rating range 2–9 mA, mean 6.2 ± 0.07 mA).

The experiment used a within-subjects A1B1A2B2 block design consisting of 2 interleaved blocks of fear conditioning in context A and extinction in context B on day 1. Each block consisted of 12 presentations of the excitatory (CS+) and inhibitory conditioned stimulus (CS−) for 3 s each. A blue and a yellow screen background served as different contexts, and 2 geometric symbols served as CSs. Thereby the CS+ in context A was in 50% of the cases paired with the aversive, individually adjusted electrotactile unconditioned stimulus (US; onset 2.5 s after CS, train of 3 square-wave pulses of 2 ms). On day 2, US intensity was individually adjusted again, followed by 10 presentations of 1 CS+ and 1 CS− in context A (acquisition context) and B (extinction context) in an interleaved design. Different contexts consisted of alternating background screen colours (blue and yellow, counterbalanced between participants). Stimulus presentation was controlled using Presentation software (NeuroBehavioural Systems).
Fear and US expectancy ratings
After every eighth trial (4 CS+ and 4 CS−, day 1) or second trial (1 CS+, 1 CS−, day 2) participants provided explicit fear and US expectancy ratings on a screen showing a picture of the CS (CS+ or CS−) within the given context, using a Likert scale ranging from 0 (none) to 100 (maximum) and answering the questions, “When you saw the symbol, how strong was your stress/fear/tension” (for fear) and “When you saw the symbol, did you expect painful stimulation?” (for expectancy).
Data recording and response definition
Skin conductance was recorded via self-adhesive Ag/AgCl electrodes placed on the palmar side of the left hand on the distal and proximal hypothenar eminence using a BIOPAC MP35 amplifier (BIOPAC Systems Inc) with AcqKnowledge 3 software. Data were down-sampled to 10 Hz, and phasic skin conductance responses (SCRs) were automatically scored as an increase in skin conductance within 1–3.5 s after CS or US onset. Specification of a minimum amplitude is optional according to published guidelines27 and was not used here. Participants not showing any US response during the first half of the experiment (day 1) were classified as nonresponders23 and excluded from all SCR analyses. Raw SCR amplitudes were logarithmized (+1) and range-corrected (SCR/SCRmax_CS).
Statistical analysis
For each dependent variable (SCRs, fear ratings, US expectancy) and day, we performed separate 2 × 2 (CS type [CS+/CS−] × context [A/B]) mixed model analyses of variance (ANOVA) with sex as the between-subjects variable. Analyses with trait anxiety, as measured by the Spielberger State–Trait Anxiety Inventory (STAI),28 as a covariate are reported in Appendix 1. For all analyses, we used appropriate follow-up tests, such as additional repeated-measures ANOVAs, for in-depth analyses of significant interaction effects. If the sphericity assumption, as tested by the Mauchly sphericity test, was violated, we adopted Greenhouse–Geisser corrections. We considered results to be significant at p < 0.05. Partial η2 is reported as a measure of effect size.
Results
Participants
In total, 377 individuals (261 women, 116 men) participated in this study. The sexes did not differ in age (range 18–35 yr, F1,375 < 1). Among the women, 172 used hormonal contraceptives and 89 were free-cycling; of the free-cycling women, 18 were in the follicular phase and 22 were in the luteal phase of their menstrual cycles.
Thirty-two participants (21 women) were classified as nonresponders23 and excluded from all SCR analyses. The SCRmax did not differ between men and women (F1,312 < 1). Groups did not differ in unconditioned SCR responses to the US (men v. women and contraceptive v. free-cycling: both F < 1; luteal phase v. follicular phase: F1,125 = 3.10, p = 0.08).
Manipulation check (main effects of task)
Generally, there was robust fear conditioning in the acquisition context A and robust extinction in the extinction context B on day 1 as well as context-dependent retrieval of fear in context A (i.e., renewal) and of extinction in context B on day 2 (Appendix 1, Fig. S2 and Fig. S3). These findings were still present in any dependent variable on any day when including sex or the use of hormonal contraceptives.
On both experimental days, there was a significant main effect of sex as well as a sex × CS type interaction in all dependent measures (fear ratings, US expectancy ratings, SCRs; Table 1 and Appendix 1, Fig. S2 and Fig. S3).
Statistical results for fear ratings, US expectancy ratings and SCRs on day 1 and day 2
Subjective ratings
On both days, there was a main effect of sex in fear and US expectancy ratings (higher in women than men) that was independent of context and CS type (Appendix 1, Fig. S2 and Fig. S3). Because this effect may reflect sex differences in the use of rating scales and because of a higher-order interaction involving the factor sex, we do not discuss the finding further. Interestingly, there was a sex × CS type interaction on both days (Table 1 and Fig. 2). This interaction was characterized by stronger differences in CS− ratings than in CS+ ratings between the sexes, resulting in attenuated CS+/CS− discrimination in women compared with men.
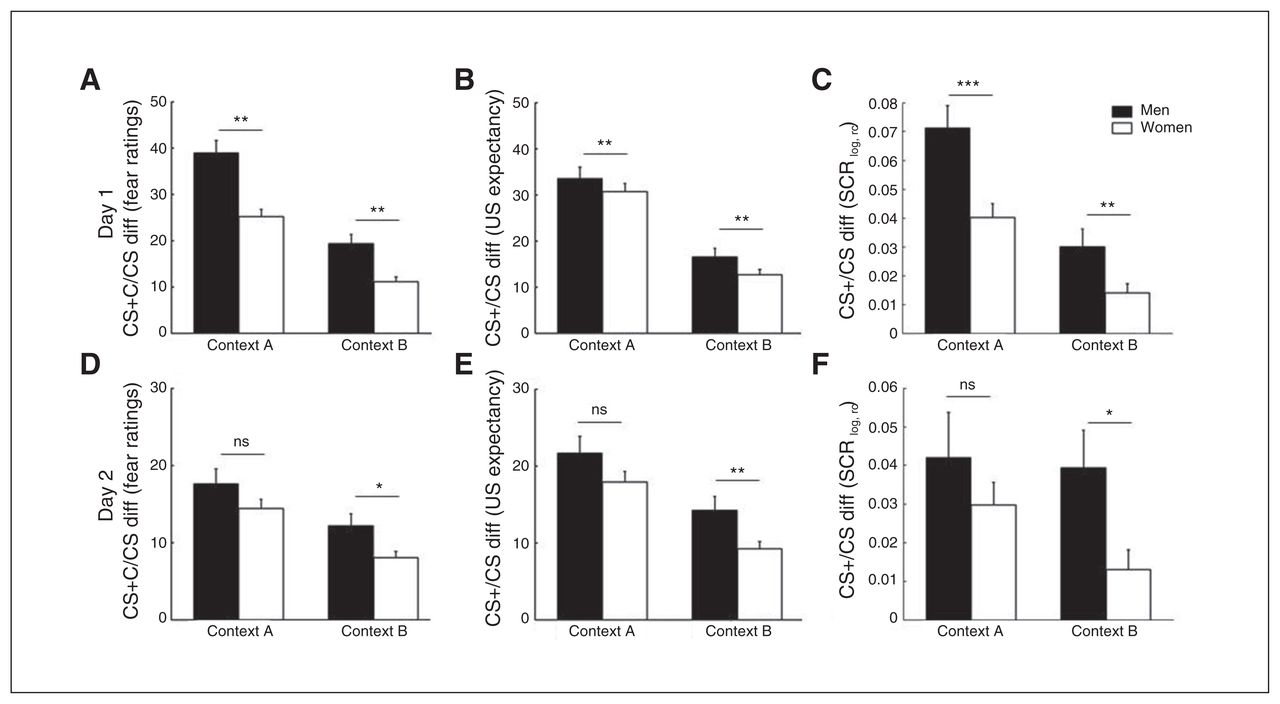
Differences in excitatory/inhibitory conditioned stimulus (CS+/CS−) scores for men (in black) and women (in white) in both the acquisition context A and the extinction context B for the 3 dependent measurements, fear ratings (A,D), unconditioned stimulus (US) expectancy (B,E) and skin conductance responses (SCRs; C,F), on day 1 (A–C) and day 2 (D–F). Error bars represent standard errors of the mean. *p < 0.05, **p < 0.01, ns = nonsignificant.
Separate analyses for both CS types showed that women had higher rating values than men, particularly for the CS− (day 1: fear ratings F1,375 = 19.17, p < 0.001, η2 = 0.05 and US expectancy F1,375 = 16.52, p < 0.001, η2 = 0.04; day 2 fear ratings F1,354 = 21.53, p < 0.001, η2 = 0.06 and US expectancy F1,354 = 19.89, p < 0.001, η2 = 0.05; Appendix 1, Fig. S2 and Fig. S3). For the CS+, the sexes did not differ on day 1 (fear ratings F1,375 = 2.54, p = 0.11 and US expectancy F1,375 = 2.00, p = 0.16; Appendix 1, Fig. S2). On day 2, however, women showed higher reactions toward the CS+ than men (fear ratings F1,354 = 11.33, p = 0.002, η2 = 0.03 and US expectancy F1,354 = 9.91, p = 0.002, η2 = 0.03; Appendix 1, Fig. S3).
In addition, on day 2 but not on day 1, the main effect of sex (fear and US expectancy in women > men) was further accompanied by an interaction with context (Table 1). Although significant in both contexts, this sex difference was more pronounced in context A (fear ratings F1,354 = 24.99, p < 0.001, η2 = 0.07 and US expectancy F1,354 = 24.11, p < 0.001, η2 = 0.06) than in context B (fear ratings F1,354 = 10.55, p = 0.001, η2 = 0.03 and US expectancy F1,354 = 9.40, p = 0.002, η2 = 0.03), giving rise to a sex × context interaction (Table 1) in the absence of a sex × CS type × context interaction.
Results were similar when excluding SCR nonresponders from rating analyses.
Skin conductance responses
For SCRs, however, men displayed larger SCRs than women on both days (Table 1 and Appendix 1, Fig. S2 and Fig. S3). This main effect of sex was further accompanied by a sex × CS type interaction, which was characterized by higher CS+ (day 1: F1,312 = 10.20, p = 0.002, η2 = 0.03; day 2: F1,310 = 5.50, p = 0.020, η2 = 0.02) but not CS− (day 1: F1,312 < 1; day 2: F1,310 < 1) reactions in men in the absence of a sex × CS type × context interaction. Hence, while reduced CS+/CS− discrimination in women (Fig. 2C and F) was observed in all dependent measures, this resulted from reduced CS+ responses in SCRs instead of higher CS− responses (as in the subjective ratings).
Effects of hormonal contraceptives
Additional analyses subdividing the women based on use of hormonal contraceptives showed no significant main effect or interaction with the use of contraceptives (yes v. no) on subjective ratings on any day and SCRs on day 1. On day 2, however, free-cycling women discriminated better than women using hormonal contraceptives in SCRs between the CS+ and the CS−, irrespective of the context (Appendix 1, Fig. S4).
Effects of menstrual cycle phase
Exploratory analyses subdividing free-cycling women further based on luteal or follicular phase of the menstrual cycle did not reveal significant differences between these sub-groups on either day (all F < 1, data not shown).
Discussion
We report several major findings. First, women displayed less CS+/CS− discrimination than men in a context-independent way on both experimental days (learning and retrieval/expression test). In particular, women showed increased CS− reactions in subjective ratings and decreased CS+ reactions in SCRs during both learning (day 1) and retrieval/expression (day 2). Second, at retrieval/expression, CS+/CS− discrimination differences in SCRs were driven by reduced discrimination in women using hormonal contraceptives compared with men and free-cycling women. Hormonal contraceptive intake and menstrual cycle phase, however, did not affect subjective ratings on any day or SCRs on day 1. Third, at the same time, women displayed generally higher fear ratings and US expectancy but smaller SCRs than men.
These findings of sexually dimorphic discrimination learning are highly important and might reflect underlying mechanisms of higher prevalence and symptom severity for anxiety and stress-related disorders in women. Discrimination between the danger (CS+) and the safety cue (CS−) is thought to represent the joint effect of 2 learning processes: excitatory danger learning (to the CS+) and inhibitory safety learning (to the CS−). Deficits in either of these processes can result in reduced discrimination rates, and an early meta-analysis on this topic implicated both mechanisms in patient–control differences in CS+/CS− discrimination.29 While in SCRs women showed reduced excitatory processing (attenuated CS+ responses), women’s subjective ratings were mainly characterized by enhanced CS− responses, likely reflecting impaired inhibitory mechanisms.30 In particular, deficient safety-signal processing represents a hallmark of anxiety-related disorders, such as panic disorder and posttraumatic stress disorder (PTSD),29,31,32 and has been implicated in theories of pathological anxiety.33 In patients wiht panic disorders, for instance, deficits to identify safety signals render panic attacks unpredictable. Ultimately, this leads to sustained anxious apprehension of the future (“better safe than sorry”).
The sexually dimorphic discrimination pattern observed in this study resembles what has been reported in patients with PTSD or panic disorder compared with controls using autonomic markers, subjective ratings and fMRI.29,32,34–39 With respect to the clinical relevance of these findings, it is tempting to speculate that impaired associative learning processes might set the ground for higher prevalence and symptom rates in women, as prospective studies have suggested impaired fear suppression as a vulnerability factor for PTSD.40,41 While the finding of reduced discrimination in this study was consistent over different dependent measures and experimental days, disparate mechanisms seem to be at work for different outcome measures (SCRs v. subjective ratings). In our data, women showed reduced autonomic responses (SCRs) to the CS+ (day 1 and day 2) and enhanced CS− responses (day 1 and day 2) in combination with enhanced CS+ responses (day 2 only) in subjective ratings.
We observed sex differences in CS+/CS− discrimination in a context-independent way, suggesting that a loss of contextual control over extinction memory, which has been implicated in psychopathology,25 does not represent a mechanism behind different prevalence rates and symptom profiles between the sexes.
Generally, literature on sex differences in fear conditioning and extinction processes is rather sparse. Still our results are well in line with those of a recent rodent study resporting faster generalization of contextual fear cues in female than in male rats,42 and the present results also fit well into a stimulus generalization framework of anxiety.43,44 Stimulus generalization represents an associative mechanism and refers to the transfer of a CR to a different stimulus that shares some characteristics (e.g., visual, physical) with the CS.45 Patients with anxiety and women may thus be more prone to a subjective initial evaluation of the CS− as a threat owing to the similarities to the true danger cue (CS+). Studies focusing on sex differences in fear generalization are ongoing, and upcoming results might clarify this. Thus, to date, it can be only speculated that women may show faster and more pronounced generalization than men.42 These results may also be of high clinical relevance not only for the understanding of the pathophysiology of anxiety disorders, but also for their treatment. If women are characterized by inhibitory learning deficits, then targeting these processes directly during therapy46,47 might be a promising avenue to precision medicine.48
Previous studies in humans have suggested activational effects of estrogens on fear conditioning and, in particular, extinction (recall) processes, as shown by differences between free-cycling women and those using hormonal contraceptives as well as differences during menstrual cycle phases.8 It was thus suggested that current sex hormone status may have a stronger impact than biological sex on fear and extinction processes. Our data stand in contrast to this and support the notion that sex differences may be more dependent on organizational effects49 as we find strong sex differences on both fear acquisition and retrieval/expression but little evidence for a modulatory role of current sex hormone status. Only during the retrieval/expression phase did we observe reduced CS+/CS− discrimination in SCRs in women using hormonal contraceptives compared with men and free-cycling women irrespective of menstrual cycle phase. This suggests a more general effect of hormonal contraceptive usage on SCRs during fear and extinction retrieval/expression rather than an effect of biological sex. In addition, no evidence for differences between subgroups of free-cycling women were observed.
Others have not observed any differences in SCRs during fear acquisition and extinction between men and women in different menstrual cycle phases either,20,21,50 but stronger CS+/CS− discrimination has been observed in women using hormonal contraceptives compared with men and free-cycling women in the luteal phase of their menstrual cycles in regions of the fear network during immediate extinction.21 In addition, women using hormonal contraceptives exhibited poorer extinction recall (i.e., stronger CS+/CS− discrimination) then free-cycling women.20 These latter findings stand in contrast to our findings of reduced CS+/CS− SCR discrimination on day 2 in women using hormonal contraceptives. Generally differences between our results and previous findings might be due to critical details in experimental designs (e.g., immediate v. delayed extinction v. extinction recall, instructed v. noninstructed acquisition, context-dependent acquisition v. cue conditioning). In addition, it needs to be acknowledged that central effects of estrogen are not mediated via serum levels but via local synthesis, even though both levels are highly correlated.51
Third, in addition to sex differences in CS+/CS− discrimination, we also observed higher subjective fear and US expectancy ratings, but generally (that is, irrespective of CS type) lower SCRs in women than in men. While it is established that women subjectively report higher fear levels,52 few studies53–55 have focused on sex differences in physiologic arousal. The significantly enhanced SCRs in men observed in our study are in line with previously reported nonsignificantly enhanced SCRs in men during fear acquisition in studies with much smaller samples than ours (Nmax = 2014,50,56). Even though SCRs were range-corrected in our study, it cannot be excluded that generally dampened electrodermal responding in females might partly contribute to differences in CS discrimination.
Limitations
While our study has several major strengths (e.g., the use of, to our knowledge, the largest sample size so far), some limitations should be acknowledged. We can provide only a rough estimate of menstrual cycle phase, as fluctuating ovarian hormones and male sex hormones were not directly measured. Consequently, classification in different menstrual cycle phases in this limited sample was based on the participants’ self-reported data. In addition, a postexperimental interview assessing CS-US contingency awareness would have been helpful to guide the interpretation of our data, as we found sex differences in US expectancy ratings and previous studies have reported differences in US expectancy and awareness between patients and controls and anxious responding.34,57,58 Furthermore, an alternative explanation to the role of biological sex might be sex-associated environmental, societal or cultural effects (e.g., sex-associated differences in upbringing; valuation of pain, fear or safety; perceived coping potential). These are, however, difficult to control in an experimental setting. Finally, up to 4 participants were tested in the same room in separate cubicles. While they could not see each other during the experiment, this might have introduced a social component that could affect results differentially for both sexes. Finally, the effect sizes for the sex differences that we observed are rather small and suggest that large samples such as ours might be required to detect them.
Conclusion
Our data demonstrate robust sex differences in CS+/CS− discrimination and safety learning and call for the incorporation of a sex difference perspective into future studies.6 Of note, it has been demonstrated previously that failure to consider sex differences may even lead to false-negative findings, such as the interaction of stress hormone levels (i.e., cortisol) with fear conditioning processes.59, 60 As estrogen is not the only sex hormone that differs between the sexes and between menstrual cycle phases, the role of other hormones (e.g., progesterone) deserve more attention, particularly in light of small effects for hormonal contraceptive usage or different menstrual cycle phases. In addition, as most rodent and human work has been conducted in males, a future challenge will be the investigation of whether results truly can be translated to women. This is of high clinical relevance given the higher prevalence and severity of anxiety symptoms in women. In light of these findings, it seems no longer appropriate to generalize experimental results obtained in men to clinical disorders that affect mainly women.
Acknowledgments
This work was supported by a State of Hamburg excellence initiative (Landesexzellenzcluster 12/09 “neurodapt”) and the Deutsche Forschungsgemeinschaft (grants KA 1623/3-1, KA 1623/4-1).
Footnotes
↵* These authors contributed equally to this work.
Competing interests: None declared.
Contributors: J. Haaker, J. Bayer, S. Brassen, N. Bunzeck, M. Gamer and R. Kalisch designed the study. J. Haaker, D. Schümann and T. Sommer acquired the data, which T. Lonsdorf, J. Haaker and R. Kalisch analyzed. T. Lonsdorf and J. Haaker wrote the article, which all authors reviewed and approved for publication.
- Received November 11, 2014.
- Revision received January 29, 2015.
- Revision received February 24, 2015.
- Accepted February 25, 2015.
References
In this issue

{kind=link}
{kind=link}
Article tools



Related Articles
Cited By...
- Fear-conditioning to unpredictable threats reveals sex differences in rat fear-potentiated startle (FPS)
- Revisiting sex differences in the acquisition and extinction of threat conditioning in humans
- Sex differences in fear regulation and reward seeking behaviors in a fear-safety-reward discrimination task
- Sex differences in fear discrimination do not manifest as differences in conditioned inhibition