

Abstract
Background: Converging evidence suggests that stress alters behavioural responses in a sex-specific manner; however, the underlying molecular mechanisms of stress remain largely unknown.
Methods: We adapted unpredictable maternal separation (UMS) and adult restraint stress (RS) paradigms to mimic stress in rats in early life or adulthood, respectively. The sexual dimorphism of the prefrontal cortex was noted, and we performed RNA sequencing (RNA-Seq) to identify specific genes or pathways responsible for sexually dimorphic responses to stress. We then performed quantitative reverse transcription polymerase chain reaction (qRT-PCR) to verify the results of RNA-Seq.
Results: Female rats exposed to either UMS or RS showed no negative effects on anxiety-like behaviours, whereas the emotional functions of the PFC were impaired markedly in stressed male rats. Leveraging differentially expressed genes (DEG) analyses, we identified sex-specific transcriptional profiles associated with stress. There were many overlapping DEGs between UMS and RS transcriptional data sets, where 1406 DEGs were associated with both biological sex and stress, while only 117 DEGs were related to stress. Notably, Uba52 and Rpl34-ps1 were the first-ranked hub gene in 1406 and 117 DEGs respectively, and Uba52 was higher than Rp134-ps1, suggesting that stress may have led to a more pronounced effect on the set of 1406 DEGs. Pathway analysis revealed that 1406 DEGs were primarily enriched in ribosomal pathway. These results were confirmed by qRT-PCR.
Limitations: Sex-specific transcriptional profiles associated with stress were identified in this study, but more in-depth experiments, such as single-cell sequencing and manipulation of male and female gene networks in vivo, are needed to verify our findings.
Conclusion: Our findings show sex-specific behavioural responses to stress and highlight sexual dimorphism at the transcriptional level, shedding light on developing sex-specific therapeutic strategies for stress-related psychiatric disorders.
Introduction
Stress is associated with increased vulnerability to many kinds of mental disorders, especially anxiety and depression. 1 As we know, biological sex is an important factor to take into consideration, as it confers differential vulnerability and resilience to stress across the lifespan.2,3 Preclinical and clinical studies have found that females and males show different responses to stress as they go through life.4–6 Converging evidence suggests that the behavioural responses to stress in males and females are complex, depending on the epochs of life, intensity of the exposure and individual responses.7 Stress during maternal pregnancy and early-life stress (ELS) put males at higher risk of proximal and some distal behavioural effects, while females have shown resilience to some adverse effects of prenatal/early postnatal stress.2 In addition, females have been shown to be significantly more resilient to adult restraint stress (RS) alone8 and combined with neonatal lipopolysaccharide (LPS) exposure.9 These animal studies suggest that males are significantly more vulnerable to ELS and adult RS, at least in terms of the measured emotional behaviours, whereas females are far more resilient.
However, the neural mechanism underlying the sexually dimorphic responses to stress remains largely unknown. Among the multiple brain areas involved in emotional and cognitive function, the prefrontal cortex (PFC) is well-known for executing top–down control and integrating diverse information about emotional processes preferentially.10 The distinguishing characteristic of the PFC is that the area is one of the last to reach full maturity in rodents and humans,11–13 and is particularly sensitive to stress exposure.14 Notably, sexual dimorphism in the PFC has been noted in human and animal studies, with remarkably earlier maturation in females than males,15,16 which may contribute to sex differences in PFC-dependent behaviours, including anxiety- and depression-like behaviours.17 Previous studies have shown that molecules in the PFC are involved in susceptibility and resilience to stress.10,18 Thus, understanding the molecular expression profiles in the PFC will help us to explain, at least in part, sexually dimorphic responses to stress.
The present study aimed to investigate the sexually dimorphic responses to stress at 2 different epochs of life. We adapted unpredictable maternal separation (UMS) and adult RS paradigms to mimic stress in early life or adulthood, respectively. Considering that stress is closely associated with increased prevalence of anxiety and depression, we assessed anxiety- and depression-like behaviours and then performed RNA sequencing (RNA-Seq) on the adult male and female medial PFC (mPFC) to identify specific genes or pathways that might be responsible for sexually dimorphic responses to stress.
Methods
Rats
Detailed information about Sprague-Dawley (Jackson Laboratories) rats and their care is provided in Appendix 1, available at www.jpn.ca/lookup/doi/10.1503/jpn.220147/tab-related-content. All procedures were carried out in accordance with the National Institutes of Health guidelines and approved by the Institutional Animal Care and Use Committee at Shanghai Jiao Tong University.
Early-life stress procedure
The UMS paradigm has been described in detail previously.19 Briefly, the separation commenced on postnatal day (PND) 4 and continued until PND 21 for on average 4 hours daily, and cages were cleaned regularly, with minimal disruption to the litter. Pups were separated from the dam and from each other. They were transferred to a room away from the dam to avoid the pups communicating with the dam via ultrasonic vocalization. After the separation period, the pups were returned to their home cage and reunited with the dam in the housing facility. Control animals were reared normally without separation from the dams except for regular cleaning of the home cages.
Adult restraint stress
Based on established RS protocol,19 our experiments used an RS paradigm (2 hours daily for 7 consecutive days) to mimic stress in adulthood briefly. Rats were individually restrained in a ventilated plastic cylinder drilled with holes to allow free breathing for 2 hours daily from PND 65 to PND 71. During the RS process, rats were immobile. After RS, rats were released and returned to their holding room with free access to food and water. The control rats were handled for 5 minutes before being returned to the holding room 2 hours later.
Behavioural procedures
All behavioural tests were conducted during the light cycle between 9:00 am and 6:00 pm under dim lighting conditions (~30 lux) from PND 72 to PND 76. The open field test (OFT) and elevated plus maze (EPM) test were administered to assess anxiety-like behaviour, and the sucrose preference test (SPT) was administered to assess anhedonia behaviour. Animals were habituated to the behavioural testing room for at least 1 hour, and the experimenter was blind to animal treatment. All data were analyzed with video tracking software (Ethovision). Further details of behavioural testing are described in Appendix 1.
RNA sequencing and differential expression analysis
Male and female rats were terminally anesthetized with isoflurane and 2 % chloralhydrate and transcardially perfused with phosphate-buffered saline (PBS) on PND 77. We then dissected the mPFC for RNA-Seq and other experiments. Total RNA was extracted using TRIzol Reagent ( Invitrogen) according to the manufacturer’s instructions. Read count normalization and gene expression estimation were performed using high-throughput sequence analysis (HTSeq). Samples were filtered for protein coding and long noncoding RNA, raw counts were summed across all samples ( Appendix 1, Table S1), and the bottom 25% were removed to eliminate genes with very low expression. The library generation protocol, sequencing parameters, quality control and subsequent analytical details can be found in Appendix 1.
Quantitative reverse transcription polymerase chain reaction
The mPFCs were disrupted and homogenized using a rotor–stator homogenizer. Total RNA from frozen dissected PFC tissue was extracted using QIAzol lysis reagent and purified using a miRNAeasy mini kit (Qiagen) according to the manufacturer’s instructions. We reverse-transcribed 400 ng of total RNA to complementary DNA (cDNA) using the High-Capacity cDNA Reverse Transcription Kit (Life Technologies). Relative expression levels of Gapdh, Uba52, Fau, Rps6, Rsl1d1l1, Rpl8, Rpl19 and Pten mRNAs were determined using quantitative reverse transcription polymerase chain reaction (qRTPCR) with SYBR Premix Ex Taq (Takara Bio), and the fluorescent SYBR Green signals were captured and analyzed with the LightCycler480 system (Roche Diagnostics). Gene expression was normalized to rat Gapdh mRNA expression. The 2−ΔΔCq method was applied for relative quantification of gene expression, and values were expressed as the mean of duplicates. Sets of specific primers are shown in Appendix 1.
Western blotting
We removed and homogenized the mPFC tissue rapidly, and we extracted proteins using a RIPA lysis buffer with 1% protease inhibitor cocktail solution (Beyotime Biotechnology). Protein concentration was detected using a BCA Protein Assay kit (Beyotime Biotechnology). Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (GenScript) and Western blotting were carried out to detect the protein levels of Uba52 (1:1000, rabbit, ABclonal), RPL19 (1:1000, rabbit, ABclonal) and β-actin (1:2000, rabbit, Cell Signalling Technology). Quantification of band intensity was analyzed using Fuji (NIMH, Bethesda MD, USA).
Statistical analysis
All statistical analyses were performed using GraphPad Prism software. We applied the normality and lognormality tests to determine the subsequent parametric or nonparametric tests. For the 2 group data sets in our study, we performed pairwise comparisons using an unpaired Student t test for normal distributions or the Mann–Whitney test for nonnormal distributions. For data with multiple measures, we analyzed main effects and interactions using 2-way analysis of variance (ANOVA). Multiple comparisons were conducted between groups if a main effect or interaction was found. Considering all the tests for both sexes were performed at the same time, all data from behavioural tests and qRT-PCR were analyzed using 2-way ANOVA, with multiple-comparison post hoc correction. All data are reported as the mean ± standard error of the mean (SEM). All statistical tests were 2-tailed and we considered results to be significant at p < 0.05.
Results
The sexually dimorphic behavioural responses to stress have been observed at 2 different epochs of life
To investigate the effects of ELS on adult males and females, we adapted a UMS protocol. We administered the OFT and EPM to assess the anxiety-like behaviour and the SPT to assess anhedonia behaviour (Figure 1A). Two-way ANOVA revealed a significant interaction effect between sex and UMS in the OFT (F1,43 = 7.202, p = 0.0103). Among male rats, time in the central zone was significantly decreased (post hoc test, p = 0.0232) after UMS, and no detectable behavioural effect of UMS was observed in females in the OFT (post hoc test, p = 0.9320; Figure 1C). There were main effects of sex (F1,33 = 19.85, p < 0.0001) and UMS (F1,33 = 21.46, p < 0.0001) on open arm ratio in the EPM. Although time spent in open arms was significantly decreased both in females (post hoc test, p = 0.0164) and males (post hoc test, p = 0.0096; Figure 1D), the decrease was more pronounced for males than females. In addition, there was a main effect of UMS (F1,41 = 11.39, p = 0.0016) on sucrose preference. Among male rats, preference for sucrose was significantly decreased (post hoc test, p = 0.0192; Figure 1E); i.e., the UMS rendered male rats more susceptible to depression-like behaviour (anhedonia in the SPT), and females did not show an elevated depression phenotype (post hoc test, p = 0.3739; Figure 1E). These data showed that UMS affected males and females differently, suggesting that male rats were more sensitive to UMS.
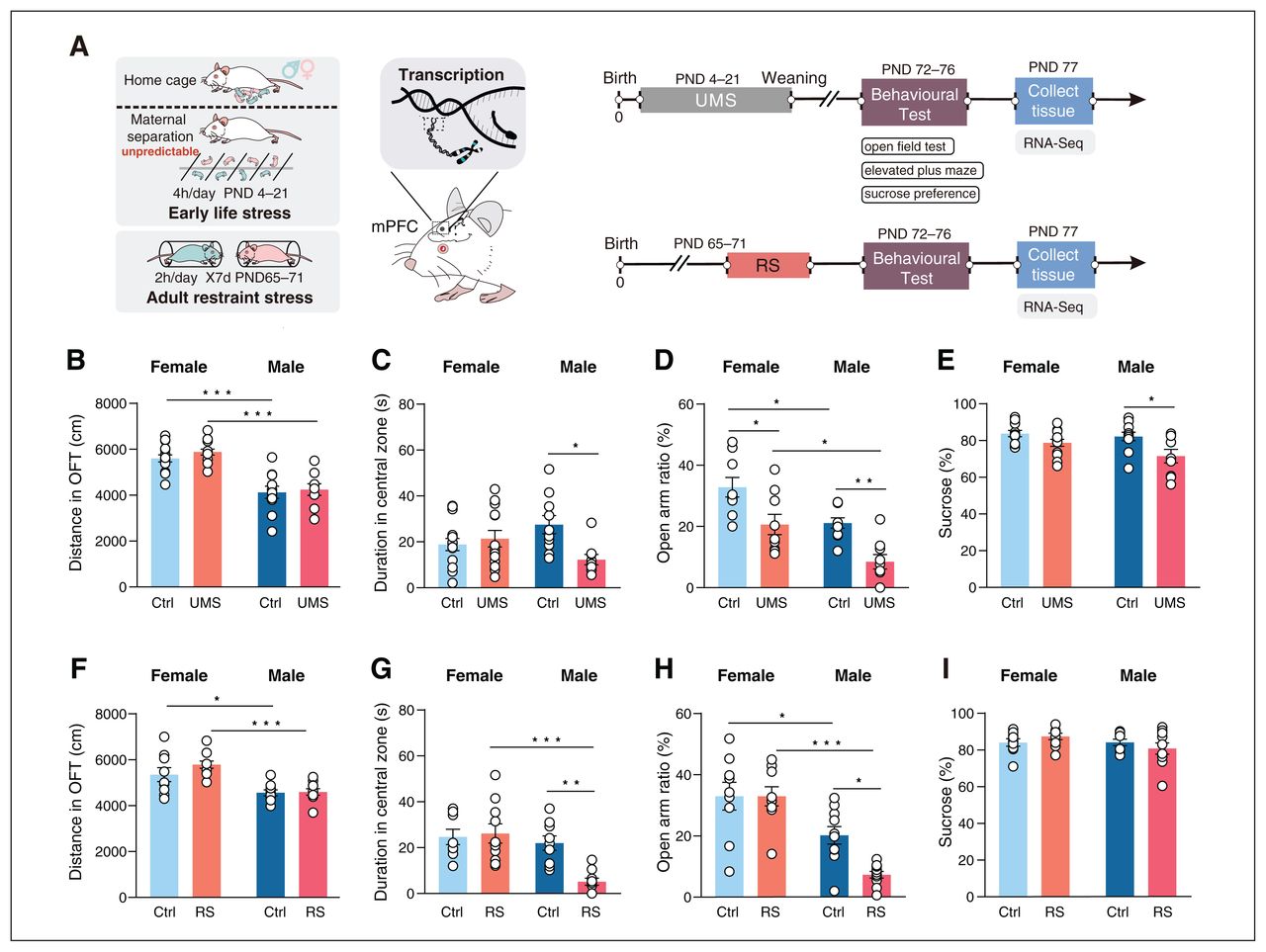
Stress has different effects on anxiety- and depression-like behavioural phenotypes in female and male rats. (A) Schematic timeline and experimental paradigm. (B–E) Comparisons of behavioural effects of unpredictable maternal separation (UMS) in female and male rats, which were affected differently. (B) Distance moved in the open field test (OFT). There was a main effect of sex on distance moved (F1,43 = 59.86, p < 0.0001). Among control rats, males showed a decreased distance moved relative to females (post hoc test, p < 0.001). (C) Duration in central zone in the OFT (2-way analysis of variance [ANOVA], interaction: F1,43 = 7.202, p = 0.0103; effect of UMS: F1,43 = 3.694, p = 0.0612; effect of sex: F1,43 = 0.004930, p = 0.9443). Among male rats, time in central zone was significantly decreased (post hoc test, p = 0.0232) after UMS, and no detectable behavioural effect of UMS was observed in females in the OFT (post hoc test, p = 0.9320). From left to right in the figure panel: n = 15, n = 13, n = 10 and n = 9 rats). (D) Open arm ratio in the elevated plus maze (EPM) test (2-way ANOVA, interaction: F1,33 = 0.008304, p = 0.92793; effect of UMS: F1,33 = 21.46, p < 0.0001; effect of sex: F1,33 = 19.85, p < 0.0001). Although time spent in open arms was significantly decreased both in females (post hoc test, p = 0.0164) and males (post hoc test, p = 0.0096), the decrease was more pronounced in males. Additionally, among control rats, males showed a decreased open arm ratio relative to females (post hoc test, p = 0.0184). From left to right in the figure panel: n = 9, n = 9, n = 10 and n = 9 rats. (E) Sucrose consumption in the sucrose preference test (SPT) (2-way ANOVA, interaction: F1,41 = 1.458, p = 0.2342; effect of UMS: F1,41 = 11.39, p = 0.0016; effect of sex: F1,41 = 3.571, p = 0.0659). Among male rats, preference for sucrose was significantly decreased (post hoc test, p = 0.0192). From left to right in the figure panel: n = 12, n = 13, n = 11 and n = 9 rats. (F–I) Comparisons of behavioural effects of adult restraint stress (RS) in female and male rats. (F) distance moved in the OFT. There was a main effect of sex on distance moved (F1,35 = 26.65, p < 0.0001). Among control rats, males showed a decreased distance moved relative to females (post hoc test, p = 0.036). (G) Duration in central zone in the OFT (2-way ANOVA, interaction: F1,35 = 8.419, p = 0.0064; effect of RS: F1,35 = 5.529, p = 0.0245; effect of sex: F1,35 = 15.66, p = 0.0004). In males, RS decreased the duration in the central zone in the OFT (post hoc test, p = 0.0033). Notably, females did not show an elevated anxiety phenotype after exposure to RS (post hoc test, p = 0.9805). From left to right in the figure panel: n = 8, n = 10, n = 9 and n = 10 rats. (H) Open arm ratio in the EPM (2-way ANOVA, interaction: F1,33 = 4.579, p = 0.0399; effect of RS: F1,33 = 4.617, p = 0.0391; effect of sex: F1,33 = 38.59, p < 0.0001). Male rats showed a decreased open arm ratio in the EPM (post hoc test, p = 0.0210) after RS. Notably, females did not show an elevated anxiety phenotype after exposure to RS (post hoc test, p > 0.9999). Additionally, among control rats, males showed a decreased open arm ratio relative to females (post hoc test, p = 0.0305). From left to right in the figure panel: n = 9, n = 9, n = 10 and n = 10 rats. (I) Sucrose consumption in the SPT (2-way ANOVA, interaction: F1,34 = 2.118, p = 0.1548; effect of RS: F1,34 = 0.00, p = 0.9972; effect of sex: F1,34 = 2.048, p = 0.1615). There was no difference in sucrose preference in male (post hoc test, p = 0.7159) and female (post hoc test, p = 0.7501) rats. From left to right in the figure panel: n = 9, n = 9, n = 10 and n = 10 rats. Ctrl = control; mPFC = medial prefrontal cortex; PND = postnatal day. *p < 0.05, **p < 0.01, ***p < 0.001.
Then we assessed the impact of adult stress on male and female rats using an established RS paradigm (Figure 1A). Two-way ANOVA revealed markedly significant main effects of sex (F1,35 = 15.66, p = 0.0004) and RS (F1,35 = 5.529, p = 0.0245) on time in the central zone in the OFT. Restraint stress decreased the duration in the central zone in the OFT (post hoc test, p = 0.0033) in male rats. Additionally, we observed a significant interaction effect between sex and RS in the OFT (F1,35 = 8.419, p = 0.0064; Figure 1G). There were main effects of sex (F1,33 = 38.59, p < 0.0001) and RS (F1,33 = 4.617, p = 0.0391) on open arm ratio in the EPM. Male rats showed a decreased open arm ratio in the EPM after RS (post hoc test, p = 0.0210; Figure 1H). Notably, female rats did not show an elevated anxiety phenotype after exposure to RS in the OFT (post hoc test, p = 0.9805; Figure 1G) or in the EPM (post hoc test, p > 0.9999; Figure 1H), suggesting that, in adulthood, females showed greater resilience to RS than males. There was no difference in sucrose preference in male (post hoc test, p = 0.7159) and female rats (post hoc test, p = 0.7501; Figure 1I), indicating that RS had no effects on depression-like behaviour. Neither UMS nor adult RS had effects on locomotor behaviour ( Figure 1B and F). Thus, our findings suggest that female rats are significantly more resilient than male rats to either UMS or adult RS, in terms of the measured anxiety-like behaviours.
Differential gene expression analysis reveals sex-specific transcriptional profiles in stressed rats
Female and male rats showed different responses to stress at 2 different epochs of life. To capture potential transcriptional patterns underlying sexually dimorphic behavioural responses to stress, we used RNA-Seq to examine the transcriptome of the mPFC in stressed rats and matched control rats. We examined 4 samples of female and male rats and found significant transcriptional changes across the mPFC in both males and females. In control rats, principal component (PC) 1 was correlated with sex (p = 8.39 × 10−7; Appendix 1, Figure 1a). We found 624 genes to be downregulated in male compared with female rats. Female rats had 1321 genes decline compared with male rats (p ≤ 0.05, false discovery rate [FDR]–corrected, fold change ≥ 2; Appendix 1, Figure 1b and Table S5). To further understand the function of differential genes, we performed enrichment analysis for differentially expressed genes (DEGs) using DAVID (2021 update). The declined genes were mainly enriched in the following pathways: cell migration (68 genes, adjusted p = 4.4 × 10−6), tube morphogenesis (68 genes, adjusted p = 4.7 × 10−5) and blood vessel development (39 genes, adjusted p = 3.1 × 10−5). The increased genes were mainly enriched in chaperone-mediated autophagy translocation complex disassembly (4 genes, adjusted p = 2.6 × 10−3) and major histocompatibility complex (MHC) class II protein complex assembly (4 genes, adjusted p = 4.6 × 10−3). Networks of enriched pathways were further analyzed using iDPE.951 (Appendix 1, Figure 1c).
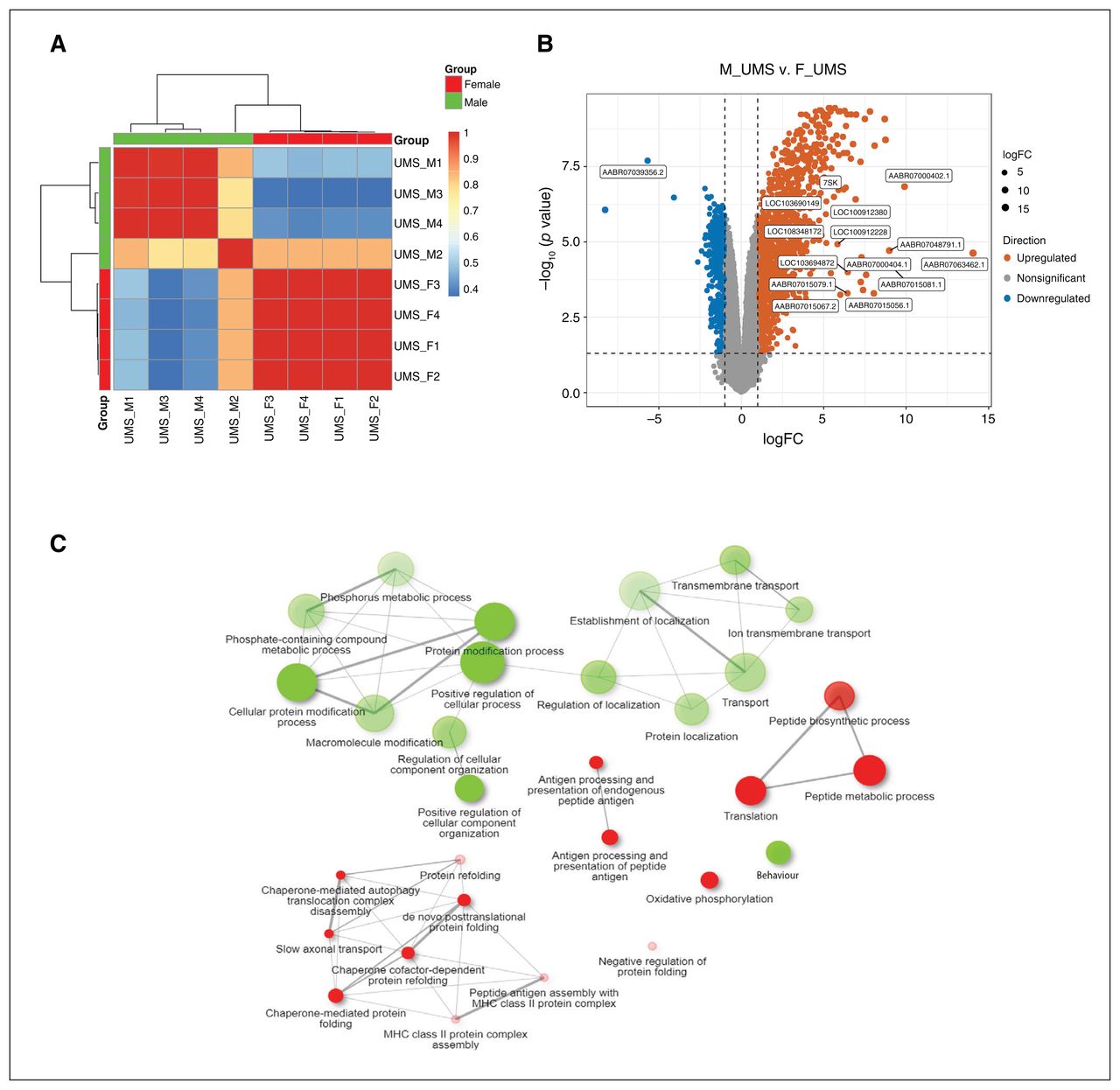
In the UMS rats, PC1 was correlated with sex (p = 2.10 × 10−9; Figure 2A). We found 759 genes to be downregulated in male rats compared with female rats. Female rats had 1380 genes decline compared with male rats (p ≤ 0.05, FDR-corrected, fold change ≥ 2; Figure 2B and Appendix 1, Table S5). The behaviour pathway (41 genes, adjusted p = 7.0 × 10−4), transport pathway (76 genes, adjusted p = 1.3 × 10−3) and protein modification process (151 genes, adjusted p = 7.0 × 10−4) were enriched in downregulated genes in male rats compared with female rats. The oxidative phosphorylation pathway (19 genes, adjusted p = 3.0 × 10−3) and chaperone-mediated protein folding (12 genes, adjusted p = 2.1 × 10−3) were enriched in upregulated genes in male rats compared with female rats (Figure 2C).

Differential gene expression analysis between female and male rats in the unpredictable maternal separation (UMS) analyses. (A) Heat map analysis. (B) Volcano plot of expression levels of the medial prefrontal cortex region in UMS rats; 759 genes were found to be downregulated in male rats compared with female rats. Female rats had 1380 genes decline compared with male rats (p ≤ 0.05, false discovery rate–corrected, fold change ≥ 2). (C) Network enrichment analysis for differentially expressed genes using DAVID (2021 update). The colour of the nodes correspond to adjusted p values. Two pathways (nodes) are connected if they share 30% (default, adjustable) or more genes. Green and red represent downregulated and upregulated pathways, respectively. Darker nodes are more significantly enriched gene sets, bigger nodes represent larger gene sets, and thicker edges represent more overlapped genes. F = female; logFC = log fold change; M = male; MHC = major histocompatibility complex.
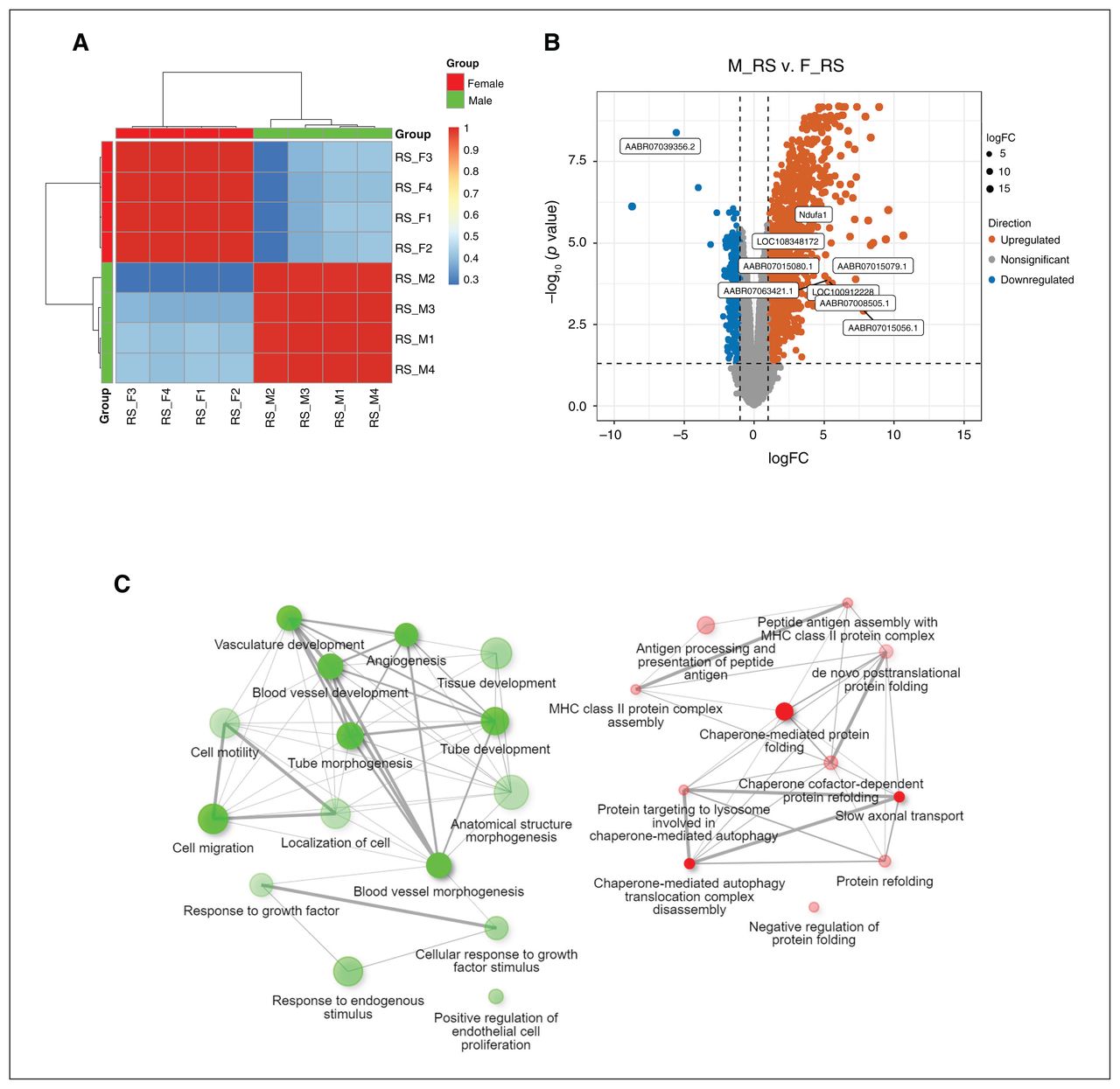
In the RS rats, PC1 was correlated with sex (p = 4.00 × 10−9; Figure 3A). We found 417 genes to be downregulated in male rats compared with female rats. Female rats had 1278 genes decline compared with male rats (p ≤ 0.05, FDR-corrected, fold change ≥ 2; Figure 3B and Appendix 1, Table S5). The blood vessel morphogenesis pathway (27 genes, adjusted p = 1.6 × 10−4), cell migration (43 genes, adjusted p = 5.4 × 10−4) and response to endogenous stimulus (43 genes, adjusted p = 1.0 × 10−3) were enriched in downregulated genes in male rats compared with female rats. Compared with female rats, the upregulated genes in male rats were enriched in chaperone- mediated protein folding (8 genes, adjusted p = 5.6 × 10−3), antigen processing and presentation of peptide antigen (13 genes, adjusted p = 5.6 × 10−3) and slow axonal transport (4 genes, adjusted p = 3.9 × 10−3; Figure 3C).
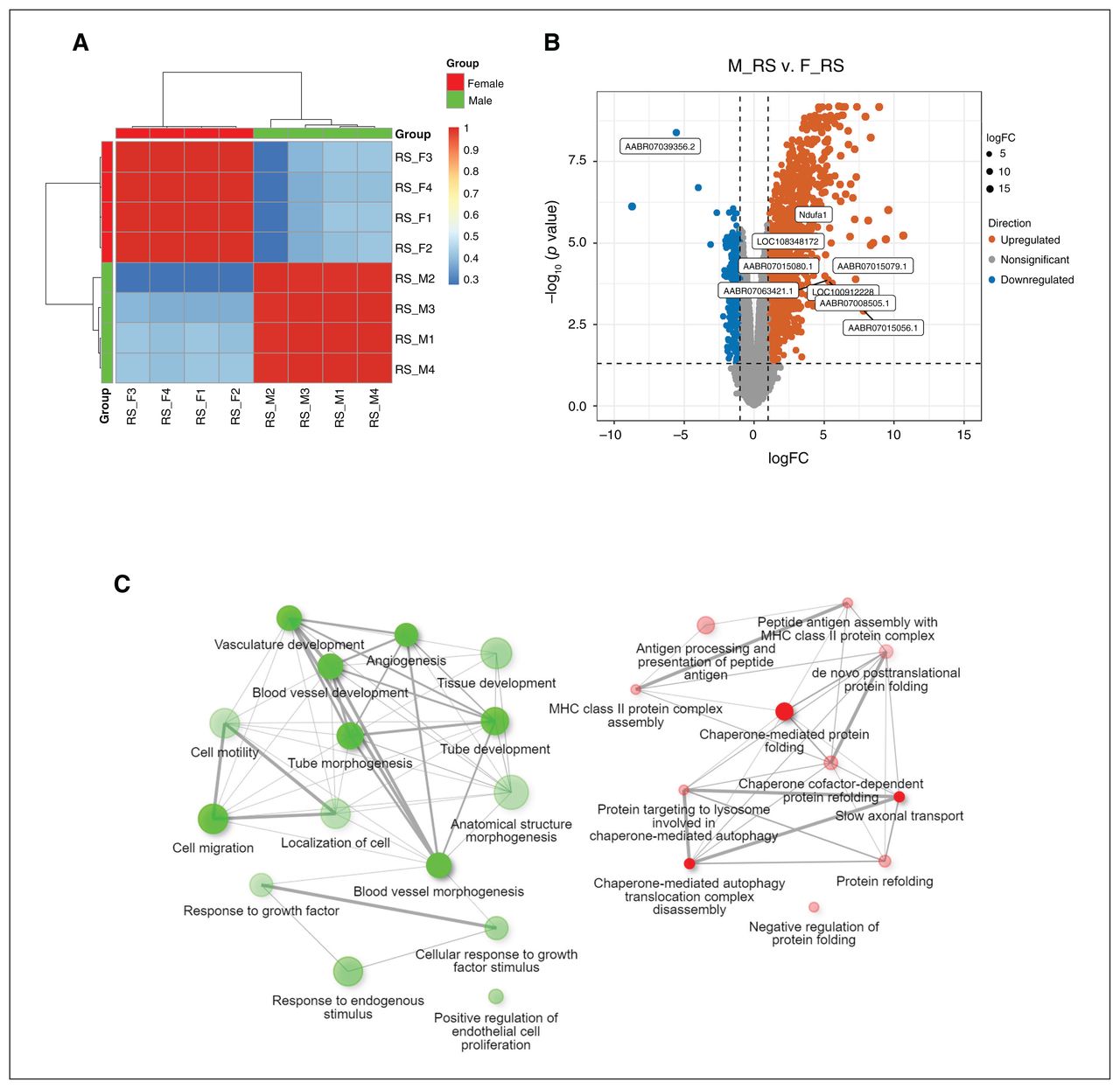
Differential gene expression analysis between female and male rats in the adult restraint stress (RS) analyses. (A) Heat map analysis. (B) Volcano plot of expression levels of the medial prefrontal cortex region in RS rats; 417 genes were found to be downregulated in male rats compared with female rats. Female rats had 1278 genes decline compared with male rats (p ≤ 0.05, false discovery rate–corrected, fold change ≥ 2). (C) Network enrichment analysis for differentially expressed genes using DAVID (2021 update). Connected gene sets share more genes. The colour of the nodes correspond to adjusted p values. Two pathways (nodes) are connected if they share 30% (default, adjustable) or more genes. Green and red represent downregulated and upregulated pathways, respectively. Darker nodes are more significantly enriched gene sets, bigger nodes represent larger gene sets, and thicker edges represent more overlapped genes. F = female; logFC = log fold change; M = male; MHC = major histocompatibility complex.
Network analyses identify sex-specific transcriptional signatures associated with stress
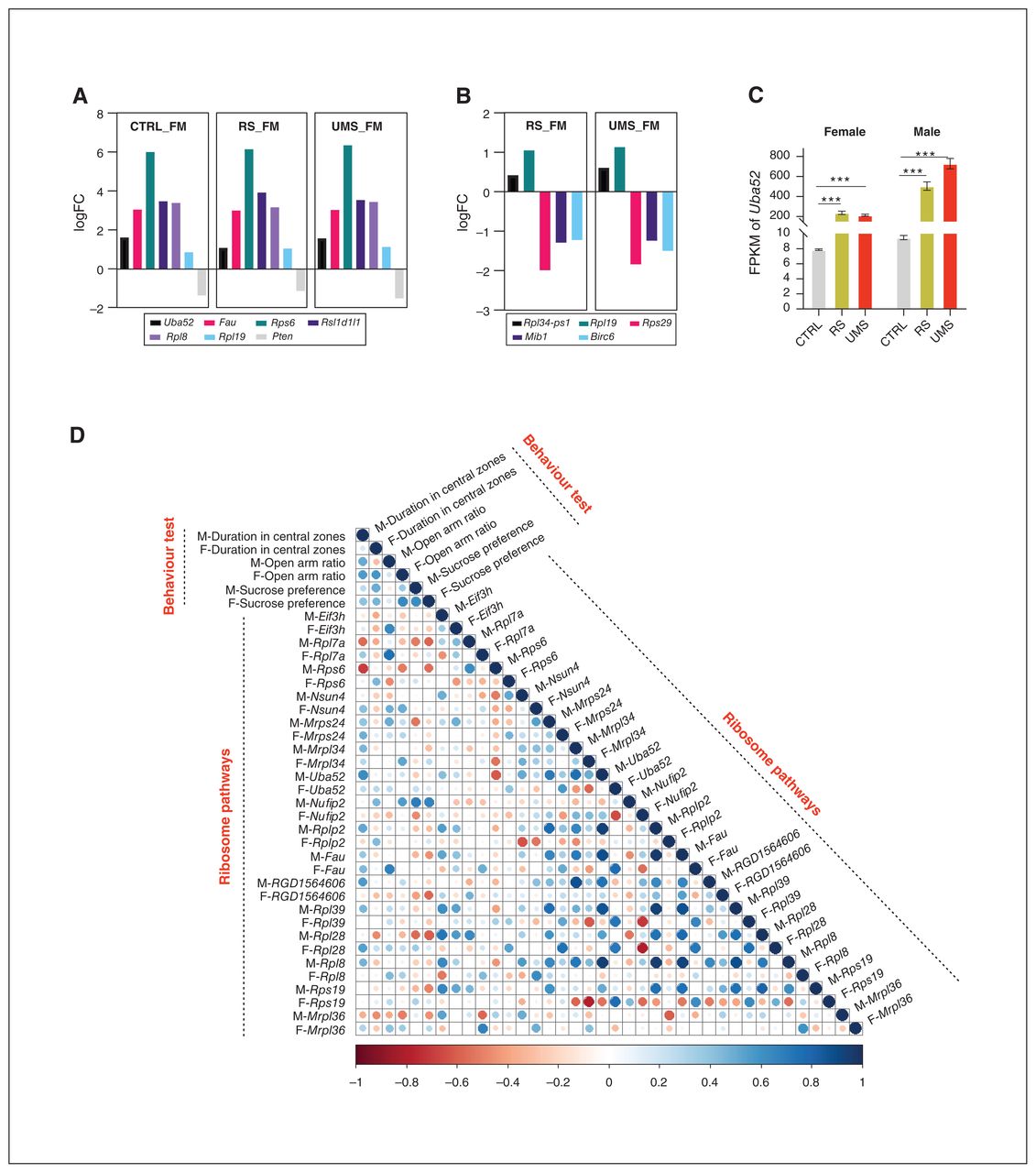
We found many overlaps in stressed transcriptional patterns between male and female rats in the mPFC. A direct comparison restricted to only DEGs in males versus females with stress revealed more than 50% overlap (Figure 4A) in the mPFC between the DEG set for UMS and RS. We found 1406 DEGs to be associated with both sex and stress, while only 117 DEGs were found to be associated with stress alone. Furthermore, we performed enriched analysis for these genes. Genes related to both sex and stress were enriched in cytosolic large ribosomal subunit, cytosolic small ribosomal subunit, ribosome, neuron projection and dendrite, while genes associated only with stress were enriched in cytoplasmic vesicle, postsynaptic density, trans-Golgi network and synapse (Figure 4A).

Network analyses identify sex-specific transcriptional signatures associated with stress. (A) Differentially expressed genes (p < 0.05) and the top associated gene ontology (GO) terms among each indicated comparison. (B) Network of 117 genes associated with stress. Green and red represent downregulated and upregulated genes, respectively. Bigger nodes represent larger gene sets. (C) Network of ranked hub gene. CTRL = control; F = female; M = male; RS = adult restraint stress; TAP = transporter associated with antigen processing; UMS = unpredictable maternal separation.
Obtaining key proteins and subnetworks in the complex differential protein interaction network are particularly important for finding the mechanism of life activities. To understand the hub genes in these DEGs, we conducted network analyses. Uba52 was the first-ranked hub gene among 1406 DEGs associated with both sex and stress (Figure 4A). Rpl34- ps1 was the first-ranked hub gene among 117 DEGs associated only with stress (Figure 4A). As shown in Figure 4C, Uba52 is of a higher level than Rp134-ps1.
To understand the differences in these hub genes between female and male rats, we listed the log fold change (logFC) in Figure 5A and B. These results showed that both in control rats and stressed rats, genes associated with ribosome pathways, including Uba52, Fau, Rps6, Rsl1d1l1, Rpl8 and Rp19, were significantly higher in male rats than in females (Figure 5A). In stressed rats, the expression of Rpl34-ps1 and Rpl19 were significantly higher in male rats than in females, while the expression of Rps29, Mib1 and Birc6 were lower in male rats (Figure 5B). The Uba52 mRNA expression is shown in Figure 5C, indicating that stress led to a sharp increase of Uba52 both in female and male rats, and male rats showed higher expression of Uba52 than females. We then analyzed the correlation between the ribosomal pathway genes and behavioural outcomes, and the results showed that the anxietylike behaviour of animals of different genders was inversely correlated with some expression of ribosome-related genes (Figure 5D). All results suggested that the ribosomal pathway was associated with sexually dimorphic behavioural responses to stress in rats.

Network analyses identify sex-specific transcriptional signatures associated with stress. (A) Fold change of hub genes (male v. female, genes associated with both sex and stress) expression in control rats and stressed rats. p < 0.05 for all listed genes. (B) Fold change of hub genes (male v. female, genes associated with stress) expression in adult restraint stress (RS) rats and unpredictable maternal separation (UMS) rats. p < 0.05 for all listed genes. (C) The expression of Uba52 in control rats, RS rats and UMS rats in RNA sequencing. (D) Associations of the behavioural test and ribosome pathways in rats. The Spearman correlation coefficient was calculated. Positive relationships are indicated in blue, and negative relationships are indicated in red. F = female; M = male.
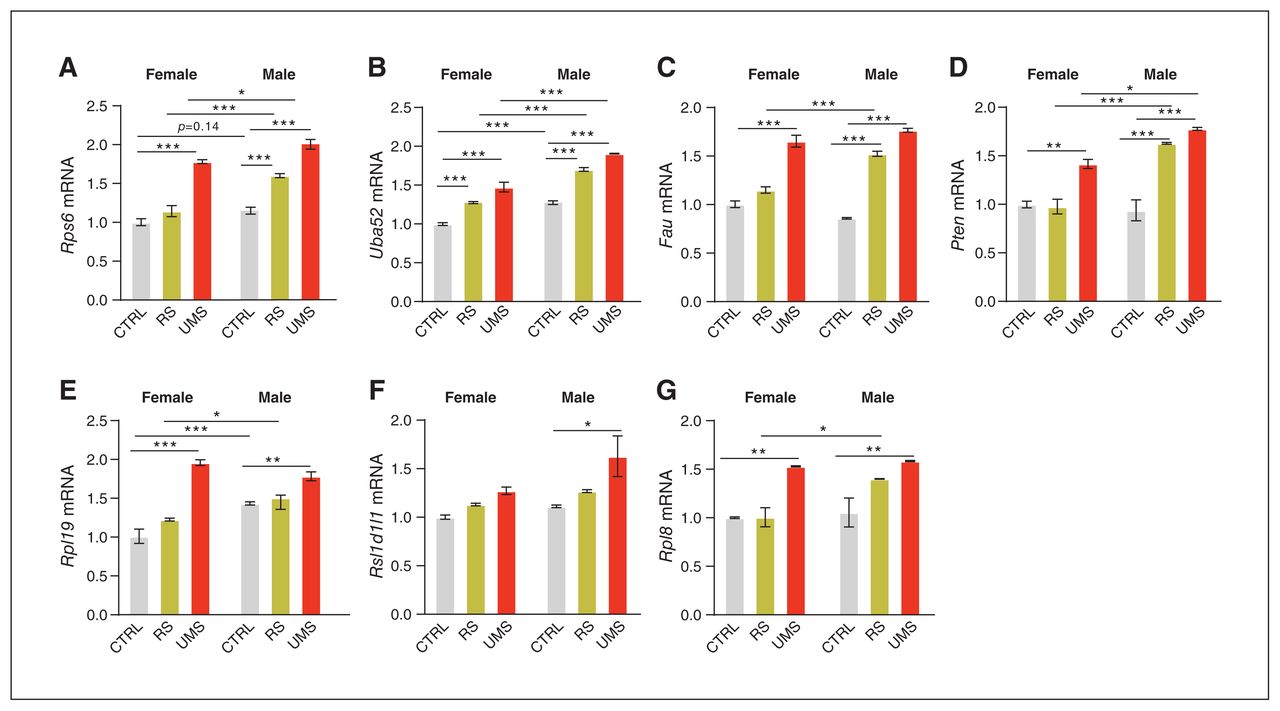
To validate the transcriptional profiles, we performed qRTPCR and Western blotting (Figure 6A–G and Appendix 1, Figure 2). For control rats, Uba52, as the first-ranked hub gene, was expressed higher in male rats than in females, which was validated by the results of qRT-PCR (p < 0.001, Bonferroni post hoc test; Figure 6B) and Western blotting ( p = 0.0495, 2-tailed unpaired t test; Appendix 1, Figure 2b). Additionally, the mRNA of Rpl19 was higher in control male rats than in females in the results of qRT-PCR (p = 0.0007, Bonferroni post hoc test; Figure 6E) and tended to be higher in Western blotting results (p = 0.0607; Appendix 1, Figure 2d). We also found that the mRNA of Rps6 was expressed higher in control male rats than in females in qRTPCR results, which is consistent with the results of RNA-Seq, although this effect did not reach statistical significance in the control condition (p = 0.14, Bonferroni post hoc test; Figure 6A). For RS rats, the mRNA of Rps6, Uba52, Fau, Pten, Rlp19 and Rpl8 was higher in male rats than in females. For UMS rats, the mRNA of Rps6, Uba52 and Pten was higher in male rats than in females. Overall, our findings suggest that ribosomal pathway genes such as Uba52 are likely associated with the sex-based difference in the observed behaviours.

Validation of the selected genes associated with both sex and stress in RNA sequencing data by quantitative reverse transcription polymerase chain reaction (qRT-PCR). (A–G) The level of Rps6, Uba52, Fau, Pten, Rpl19, Rsl1d1l1 and Rpl8 mRNA expression in the medial prefrontal cortex. All data from the qRT-PCR were analyzed using 2-way analysis of variance, with Bonferroni post hoc multiple comparisons correction. CTRL = control; F = female; M = male; RS = adult restraint stress; UMS = unpredictable maternal separation.
Discussion
To our knowledge, this work is the first to directly show the sexually dimorphic responses to stress during early life and adulthood. To understand the molecular mechanisms of sex differences, underlying transcriptional patterning in the mPFC was further explored. Our findings of how stress differentially affected male and female rats provides new insight into stress-related mental illness.
Consistent with previous reports on the stress resistance of females in many domains,4,5,8,20 our study provides direct evidence that females were significantly more resilient to either UMS or RS in terms of the measured anxiety-like behaviours. However, epidemiological studies have indicated that the prevalence of stress-related psychiatric disorders, such as depression and posttraumatic stress disorder (PTSD), is greater among women than men. This has led some to conclude that women may have higher stress susceptibility, but it is tough to identify the biological or social factors responsible for the generation of these sex differences. Social factors may play a bigger role than biological sex.21 Generally, women tend to experience greater social inequality and social stress. As shown in a study investigating stress in more than 2500 health care providers on the front lines during the COVID-19 pandemic, although women were more likely than men to report symptoms of psychological distress (42% v. 30%), gender differences were not found when taking into account pre- existing and concurrent stressors, which are more often experienced by women.22 Consistently, Assari and Lankarani23 found that the effect of each life stressor on the risk of clinical depression was 50% stronger for men than women on their rates of depression 25 years later, which corresponds with their findings from another study showing that white men may be most vulnerable to the effect of stress on depression.24 In addition, genetic factors carried by susceptible individuals are prone to influence the intrinsic sensitivity to stress directly, and stress may act only as a trigger to provoke emotional difficulties.25
Considering confounding factors related to social psychology and genetic risk in human studies, animal studies are a better option for exploring the biological sexually dimorphic responses to stress and underlying neural mechanisms. Notably, the observed sex differences in stress responses in animal studies could be affected by the timing of the exposure, and there appears to be some disparity in the conclusions among different animal studies.6,7 Unlike in studies involving mice, there are not many widely used paradigms in studies involving rats. Maternal stress26,27 and RS4,8,28 have been the typical stress paradigms used in rats during early life and adulthood, respectively, and they have been adapted widely to observe sex differences in the impact of stress. Thus, we adapted 2 typical stress paradigms (UMS and RS) in early life and adulthood, respectively, to assess sexually dimorphic responses to stress. Additionally, our 7-day RS model was adapted based on previous literature4,8 and our preliminary experiments. Wei and colleagues4 found that female rats exposed to 1 week of RS for 2 hours daily showed no negative effects on temporal order recognition memory, which induced the impairment in males.4 Another study showed that acute RS (a single 2-hour exposure) led to higher anxiety-like behaviour in male rats, whereas females did not show delayed increase in anxiety-like behaviour.8 These findings suggest that females and males show different responses to RS of even shorter duration than 7 days, thus making RS a robust model for follow-up research into molecular mechanisms underlying sex differences.
Prior studies have noted profound sex differences in transcriptional responses to stress during both early life29,30 and adulthood.31 Here, leveraging DEG analysis of RNA-Seq in the mPFC, we highlight the fundamental transcriptional differences in male and female rats under either UMS or RS, and our findings were consistent with those of previous studies to some extent. Notably, our findings differed from those of other studies in that we depicted the overlaps of DEGs affected by both ELS and adult stress and identified a potential pathway in the mPFC likely involved in stress-induced pathology. Among these overlaps, DEGs were mostly found to be associated with both biological sex and stress and were mainly enriched in the ribosomal pathway. There is increasing evidence that the ribosome is linked to the cellular responses to external and internal environmental stimuli.32 As we know, ribosomes are essential subcellular organelles where biological protein synthesis takes place. The ribosomal proteins (RPs) of the large (RPL) subunit and small (RPS) subunit — the 2 components of the ribosome — are critical for ribosome assembly and function. The process of ribosome biogenesis is tightly regulated and highly dynamic, modulated by multiple signalling pathways responding to growth factors, energy and stress.33,34 Defects in either the structure or function of these ribosomal components (ribosomopathies) have been well documented in metabolic and hematological diseases as well as cancer. In addition, ribosomes are known to contribute to the development of neurons in the central nervous system, and the lowered ribosomal activity, reduced protein synthesis and consequent growth arrest have been observed under stress situations.32,35,36
Ribosomal genes, particularly RPL17 and RPL34, as key molecules play an integral part in stress vulnerability across nonclinical and clinical conditions.37 Notably, the dysfunction of RPs has been observed in the progression of stress-related psychiatric disorders in humans, including PTSD and depression. Differential expression in the ribosomal pathway has been observed in the blood samples of cohorts with PTSD compared with controls.38 Weighted gene coexpression network analysis identified that the functional module of ribosome biogenesis was strongly correlated with the development of major depression.39 Consistently, another study found that the upregulation of the ribosomal pathway was a potential blood biomarker for comorbid major depressive disorder and PTSD.40 In rodents, the imbalanced regulation of ribosome-related genes in the PFC in males has been linked to stress susceptibility.41 In addition, researchers have performed RNA-Seq in male mouse brain, exposing chronic social defeat stress, and have reported ribosomal dysfunction in genes including RPL6 and RPL29, which contributes to anxiety- and depression-like symptoms.42 These findings indicate that dysfunction of the ribosomal pathway contributes to the pathological mechanism of stress-related psychiatric disorders. Indeed, in our study the observed behavioural changes after stress were likely mediated partially by the ribosomal pathway. The correlation data strongly support this idea. Perturbations in ribosome biogenesis and function may activate p53- mediated nucleolar stress response involving multiple RPL and RPS and further transactivate downstream targets, leading to senescence, cell cycle arrest, apoptosis or autophagy.35 Thus, in the central nervous system, ribosome biogenesis plays a vital part in regulating neural cell growth and proliferation; any dysregulation of this process may result in higher risk of developing stress-related psychiatric disorders and may contribute to male rodents’ vulnerability to stress.
An important finding in our study is that ribosomal signalling was linked to biological sex. Uba52, as the first-ranked hub gene among the set of 1406 DEGs, was more highly expressed in male rats than females under either the control or stress conditions. Uba52 encodes a ubiquitin-ribosomal fusion protein consisting of ubiquitin at the N-terminal region and ribosomal protein L40 (RPL40) at the C-terminal region, which is a main source of ubiquitin protein.43 Ubiquitin is a critical signal protein regulating many cellular processes by modulating protein abundance, and the balance of ubiquitin pools is deeply coupled to various stressors, which is associated with several neurologic disorders.44 Importantly, increased expression of ubiquitin has been observed following exposure to stressors including oxidative stress, heat, ischemia and infection.45,46 Chronic overexpression of ubiquitin in neurons impairs cognitive function and synaptic plasticity in mice.47 Interestingly, in our study, male rats showed a more highly increased expression of Uba52 than females under the RS and UMS paradigms, although a trend toward upregulated Uba52 has been detected in both males and females. In addition, Uba52 was significantly associated with the observed behaviours, highlighting regulatory effects of the ribosomal pathway on sex-specific stress susceptibility. Consistently, Devulapalli and colleagues48 found that male rats showed increased freezing time compared with females in a conditioned fear paradigm and reported that sex differences in the regulation of ubiquitin-proteasome mediated by Uba52 in the amygdala likely underlie the sex-related behavioural differences in fear memory formation. Furthermore, genetic suppression of Uba52 expression in the amygdala reduced freezing time in male rats, which supports the important role of Uba52 in the stress-induced behavioural phenotype. 48 Thus, herein we propose that the excessive elevation of Uba52 impairing neuronal function and behaviour renders male rats more susceptible to stress than females. Uba52 has been found to modulate the ribosomal protein complex dually, contributing to the biogenesis and function of ribosomes, 49 indicating that ribosomal signalling is more sensitive to stress in male rats than in females. Furthermore, sex differences in ribosomal signalling have also been observed in ischemic stroke–induced peripheral immunosuppression. 50 These data may suggest that the dysfunction of ribosomes and corresponding cellular signalling likely underlie sex disparities in behavioural responses to stress.
It is important to note that Uba52 was more highly associated with both biological sex and stress than Rp134-ps1 was associated with only stress. Uba52 encodes RPL40 and ubiquitin for the ribosome simultaneously, modulating stress response. Rp134-ps1 encodes a protein likely involved in the synthesis of cytosolic RPL. We can further infer that stress may lead to more pronounced effect on the set of DEGs related to both sex and stress enriched in the ribosomal pathway. The DEGs associated with only stress were primarily enriched in synapse-related pathways. Notably, 1 study reported that ribosomes constitutively supply synapses with proteins51 in the rodent brain, and continued mRNA translation and protein production of ribosomes are essential for neuronal synaptic activity52,53 and homeostasis,54 suggesting that ribosomes affect synaptic structure and function in the central nervous system. The positive correlation between the number of ribosomes and the thickness of postsynaptic density has been observed in dendrite terminals of neurons,55,56 providing evidence for the importance of ribosomes in ultrastructural synaptic plasticity. Consistently, neuronal ribosomal protein mRNAs have been frequently detected in dendrites and axons, which is crucial for synaptic modifications like long-term depression and long-term potentiation.54 Additionally, presynaptic protein synthesis in ribosomes is critical for maintaining the release of neurotransmitters during long-term plasticity.57,58 Taken together, it seems more likely that the activation of protein synthesis in ribosomes necessary for synaptic plasticity may partly uncover the molecular mechanisms underlying sex-specific responses to stress susceptibility in the PFC.
Limitations
Sex-specific transcriptional profiles associated with stress were identified in this study, but more in-depth experiments, such as integrating single-cell approaches to identify cell-type–specific molecules and gene regulatory networks and performing manipulation of male and female gene networks to validate our bioinformatics predictions in vivo, are needed to verify our findings. In addition, our study detected sex differences of stress responses and neural mechanisms in the PFC of rodents; confounding factors related to social psychology and genetic risk in humans require consideration.
Conclusion
We identified and verified sexually dimorphic responses to stress in rats at 2 different epochs of life, and ribosomal pathways may underlie sex-specific behaviours in association with susceptibility to stress. Future molecular studies should validate and extend this work to identify the specific mechanism of ribosomes in vitro and in vivo. Overall, our study provides evidence of the molecular and cellular mechanisms underlying sex-specific stress susceptibility, which may inform the development of sex-specific therapeutic strategies for stress-related psychiatric disorders linked to deregulated ribosomal pathways.
Acknowledgements
This work was supported by grants from the Shanghai Science and Technology Committee (20XD1423100, 19YF14420000), the Shanghai Municipal Education Commission (2021-01-07-00-02-E0086), the Xuhui District Artificial Intelligence Medical Hospital Cooperation Project (2021-005) and the National Natural Science Foundation of China (82230045, 32271066, 31900730).
Footnotes
↵* Contributed equally to this study.
Contributors: Y.-D. Zhang, D.-D. Shi and Z. Wang designed the study. Y.-D. Zhang, D.-D.- Shi and S. Zhang acquired the data, which all authors analyzed. Y.-D. Zhang and D.-D. Shi wrote the article, which all authors reviewed. All authors approved the final version to be published, agreed to be accountable for all aspects of the work and can certify that no other individuals not listed as authors have made substantial contributions to the paper.
Competing interests: None declared.
- Received August 24, 2022.
- Revision received November 3, 2022.
- Revision received November 18, 2022.
- Accepted November 18, 2022.
This is an Open Access article distributed in accordance with the terms of the Creative Commons Attribution (CC BY-NC-ND 4.0) licence, which permits use, distribution and reproduction in any medium, provided that the original publication is properly cited, the use is noncommercial (i.e., research or educational use), and no modifications or adaptations are made. See: https://creativecommons.org/licenses/by-nc-nd/4.0/
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Article tools



Related Articles
Cited By...
- No citing articles found.