

Abstract
Environment is known to substantially alter mental state and behaviour across the lifespan. Biological barriers such as the blood–brain barrier (BBB) and gut barrier (GB) are major hubs for communication of environmental information. Alterations in the structural, social and motor environment at different stages of life can influence function of the BBB and GB and their integrity to exert behavioural consequences. Importantly, each of these environmental components is associated with a distinct immune profile, glucocorticoid response and gut microbiome composition, creating unique effects on the BBB and GB. These barrier–environment interactions are sensitive to change throughout life, and positive or negative alterations at critical stages of development can exert long-lasting cognitive and behavioural consequences. Furthermore, because loss of barrier integrity is implicated in pathogenesis of mental disorders, the pathways of environmental influence represent important areas for understanding these diseases. Positive environments can be protective against stress- and age-related damage, raising the possibility of novel pharmacological targets. This review summarizes known mechanisms of environmental influence — such as social interactions, structural complexity and physical exercise — on barrier composition, morphology and development, and considers the outcomes and implications of these interactions in the context of psychiatric disorders.
Environment, barriers and the brain
Every organism is shaped by their surroundings through constant perception, consumption and physical engagement with environmental features such as chemical makeup, social targets, structural complexity and climate.1,2 All of these environmental components interact with the genome to influence phenotype and, thus, cognitive ability and behaviour, throughout development and aging. The idea that the environment could permanently affect the adult brain was popularized by Donald O. Hebb, who found that rats living in his home as pets performed much better on memory tasks than those raised in laboratory cages.3 Later work, especially from the laboratory of Mark Rosensweig, was crucial in deciphering the neurobiological underpinnings of this phenomenon; 4,5 today, we know that environmental conditions affect both neuronal plasticity and functional connectivity, as well as genesis of new neurons, glia and cerebral blood vessels.6–10 But the question remains, how exactly is external information translated into lasting effects on the brain? Although activity-dependent changes in neuronal plasticity offer an enticing and simplistic mechanism, evidence increasingly supports a role for changes to the periphery including the immune system, microbiome and glucocorticoid responses, which occur independently from the brain and yet are indispensable for some of the cognitive and behavioural consequences of environmental change.11–16 These systemic pathways for brain–environment communication, which remain poorly characterized, rely heavily on communication across specialized biological barriers, including the blood–brain barrier (BBB) and gut barrier (GB).
The BBB and GB are conserved, highly specialized frontiers that protect the body from external disturbances while maintaining communication between anatomic compartments. The BBB, a dynamic endothelial membrane lining the lumen of blood vessels in the brain, is essential to cerebral homeostasis, supplying necessary nutrients and metabolites to neurons and glia while preventing entry of harmful substances from the bloodstream.17 The BBB is supported by an intricate cell network surrounding cerebral blood vessels termed the neurovascular unit, mainly comprising astrocytes, mural cells (including smooth muscle cells and pericytes), microglia and brain endothelial cells (Figure 1).18–20 The neurovascular unit performs many crucial functions including regulating BBB development and maintenance, and facilitating coupling between neuronal activity and cerebral blood flow.21 Specific BBB properties are largely conferred by brain endothelial cells that possess several unique features, including a tightly regulated transporter profile, restricted expression of adhesion molecules and a specialized tight junction architecture between neighbouring cells (Figure 1).17,22 Importantly, this composition is highly sensitive to circulating signals from the periphery, including cytokines and immune cells, glucocorticoid levels and gut-derived bacterial metabolites.23
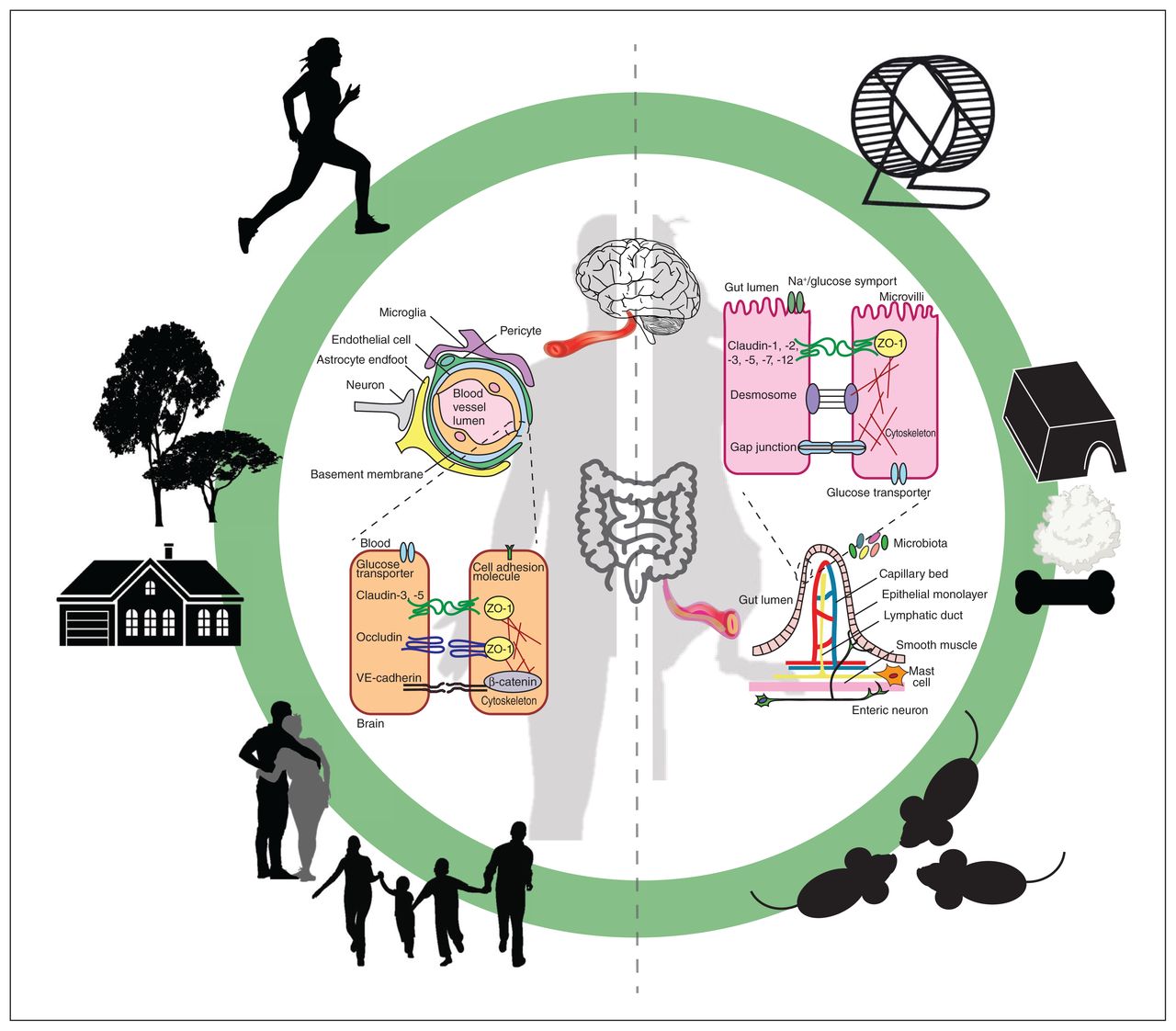
Biological barriers are key interfaces between environment and the body. The blood–brain barrier (BBB, inner left) is a dynamic and highly selective membrane between the endothelial cells that line the blood vessels of the brain. Specific barrier properties are conferred by specialized tight junction proteins (e.g., claudin-5) and by occludins anchored to the cytoskeleton by scaffold proteins, including zonula occludens (ZO). The BBB is encircled by an intricate network of cells termed the neurovascular unit, mainly comprising the following: astrocytes, which facilitate neuronal communication with blood vessels; mural cells including smooth muscle cells (on arteries and arterioles) and pericytes, which contribute to contractile properties and morphology of vessels; and microglia, the resident immune cells of the brain. Meanwhile, the gut barrier (GB, inner right) is a layer of epithelial cells termed enterocytes lining the lumen of the intestine. These cells are connected by a unique network of tight junctions — particularly claudins, as well as desmosomes and gap junctions — that regulate passage of substances from the gut lumen to the body. The GB is in close contact with the intestinal microbiome and is sensitive to changes in bacterial function. In addition, the GB communicates with enteric neurons, blood vessels, mast cells and immune cells, making it a key signalling hub. Both of these barriers mediate communication between the environment and the body in humans and in rodent models. Distinct environmental components — such as social interactions, structural complexity and locomotor activity — which can be translated from rodents (outer right) to humans (outer left), influence the barriers in specific ways to produce unique physiologic effects. VE = vascular endothelial.
The GB — an epithelial membrane embedded in the mucosal layer of the gut lumen — plays a crucial role in homeostasis of the gastrointestinal tract. Given its close contact with the external environment, the GB is a converging centre for interactions between diet, microbiome, immune system and enteric efferent neurons (Figure 1).24,25 A brick-like tight junction pattern along the extended site of contact between gut epithelial cells closely regulates passage of fluids and metabolites from the gut lumen to the bloodstream (Figure 1).26 The GB interacts physically and chemically with intestinal microflora, which are sensitive to environmental changes and communicate with the body through metabolite levels, such as short-chain fatty acids (SCFAs), as well as microbial surface proteins, including peptidoglycans and lipopolysaccharide.27,28 Produced in varying amounts by specific bacterial strains, SCFAs and lipopolysaccharide are key messengers that influence cognition and behaviour through a communication network known as the microbiota–gut–brain axis.28,29 Despite their differences, the BBB and GB are both integral to survival throughout the lifespan. Indeed, each is required for normal embryonic development, beginning at early stages; loss of barrier integrity in later life is associated with cognitive and physiologic deficits of aging.30–34 Cooperation and communication between the BBB and the GB are therefore important aspects of normal physiology, mediated by a constant exchange of peripheral signals in response to environmental demands.
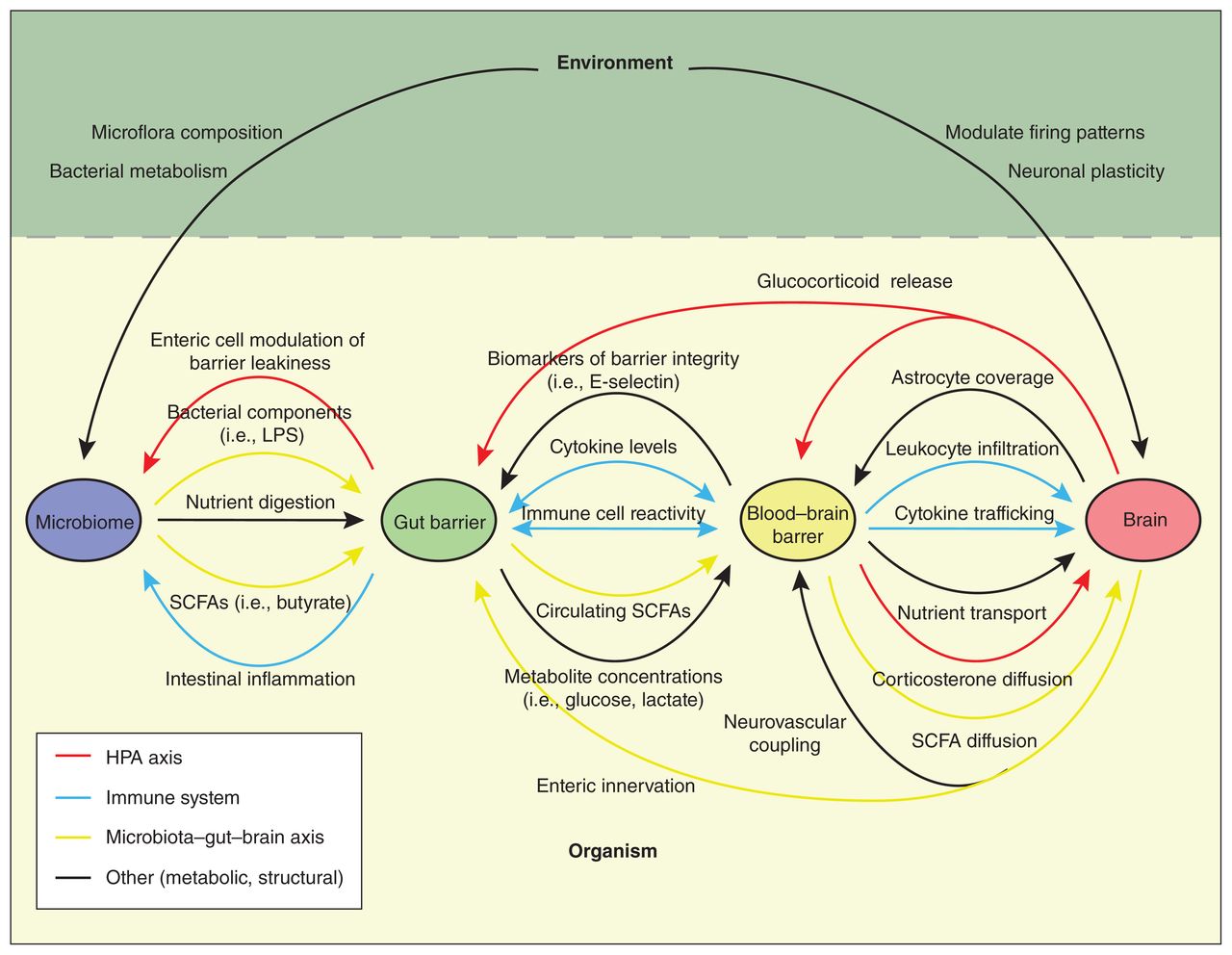
Much of the environmental influence on cognition and behaviour throughout life is dependent on translation of external signals to the brain via the periphery. Importantly, the BBB and GB are not simple mediators of this communication, but rather active signalling hubs that sense and respond to relevant information through changes in receptor profile, gene expression and barrier permeability (Figure 2). The peripheral immune system is a major pathway for these interactions, as levels of circulating cytokines and leukocytes are highly sensitive to surrounding environment and fluctuate with conditions such as temperature and air pollution. 11,35 These produce a variety of barrier effects; for example, elevated levels of proinflammatory cytokines like tumour necrosis factor-α (TNF-α) and interleukin-6 (IL-6) suppress transcription of tight junction proteins and diminish BBB and GB integrity to allow infiltration of inflammatory mediators into the bloodstream and the brain.36–38 The microbiota–gut–brain axis provides an alternative avenue for interaction, where microbial metabolites such as SCFAs, which vary with environmental conditions, can influence intestinal barrier permeability, as well as diffuse across the GB into the circulation to influence leukocyte count, BBB integrity and neuronal activity.28,39 Meanwhile, lipopolysaccharide leaks into the bloodstream via a paracellular route between epithelial cells, where it forms a complex with circulating soluble binding proteins (e.g., lipopolysaccharide binding protein), which can alter the BBB and cognitive function.27,36 Finally, the physiologic stress response via the hypothalamic–pituitary–adrenal (HPA) axis communicates with and across barriers through levels of circulating glucocorticoids. When perceived stress initiates HPA activation, steroids — such as cortisol (in humans) or corticosterone (in rodents) — are released into the blood, where they interact with barriers through their binding with 2 kinds of receptors, the mineralocorticoid and glucocorticoid receptors.40,41 The binding of these steroids in the barrier modifies their permeability and responsiveness to subsequent stimuli.42,43

Pathways of environment–barrier communication. The blood–brain barrier (BBB) and gut barrier (GB) communicate with each other and with the environment through a complex network of interactions. The main avenues for signalling are the hypothalamic–pituitary–adrenal (HPA) axis (red), the immune system (blue) and the microbiota–gut–brain axis (yellow). Information received from the environment produces changes in neuronal activity or microbiome function that are sensed by the GB and BBB and are subsequently transduced into circulating signals, enabling the brain to be affected by shifts in the gut and vice versa. LPS = lipopolysaccharide, SCFA = short-chain fatty acid.
These interactions take on a special importance in the context of disease, as growing evidence suggests that disruption of environment–barrier communication is an early step in pathogenesis of mood disorders. Several lines of evidence have recently come together to support this notion. Disruption of the BBB — measured with magnetic resonance imaging by injection of contrasting agents or in postmortem tissue with markers of BBB integrity — and the GB — inferred from blood serum markers of GB permeability — have been observed in both males and females with diverse psychiatric conditions including major depressive disorder (MDD), bipolar disorder and schizophrenia.36,38,44–47 In mice, chronic social or psychological stress is sufficient to induce depressive and anxiety-like symptoms via damage to the BBB and GB (Doney and colleagues25 provide a review of BBB and GB disruption in various rodent models of stress).36,38,48–50 Environmental stressors such as social isolation, low socioeconomic status or sedentary lifestyle have been associated with systemic inflammation and risk for developing psychiatric disorders in both rodents and humans.51–55 Meanwhile, positive environments can protect against disease; in humans, physical exercise, high socioeconomic status and social support decrease risk for psychiatric disorders,54,56,57 while enriched housing or access to a running wheel have strong antidepressant effects in mouse models of stress.58,59 Finally, several reports have suggested that the positive behavioural outcomes of enrichment could involve changes to the neurovasculature and gut epithelium.8,60–64 As current monoamine-focused antidepressant and anxiolytic medications are, unfortunately, not always effective,65 positive environmental modifications to the BBB and GB could offer new and attractive pharmacological targets for mood-related disorders.
Although the BBB and GB are increasingly recognized for their major role in environmental influence on cognition, behaviour and emotion in health and disease, the literature on this subject is broad, confusing and, at times, contradictory. Therefore, we aim to summarize known mechanisms of environmental influence on barrier activity, morphology and development, as well as to consider the outcomes and implications of these interactions in the context of psychiatric disorders to identify directions for future research.
Distinct environmental features differentially modulate biological barriers
An organism’s environment is extraordinarily complex, with many diverse aspects that can combine or cancel out as they interact with the genome to shape behaviour and cognitive function. Investigation of these relationships in a laboratory setting must strike a balance between feasibility and ethological validity. The most common means of studying neural consequences of various external conditions in the laboratory is the addition of extra factors to the home cage, a paradigm generally referred to as environmental enrichment.2,5 However, what enrichment actually means has been subject to various interpretations over the years. Cage layouts in these experiments tend to vary greatly among laboratories, and the literature is replete with a heterogeneous mix of enrichment paradigms including larger cages, multiple levels, cage mates, running wheels, shelters, toys, enhanced diet and added bedding, often combined.66 The consensus definition of enrichment as “a combination of complex inanimate and social stimulation”2 suggests an interweaving of factors that are not easily separated. However, it is increasingly apparent that distinct environmental components have unique effects on the brain and, therefore, their effects should be studied independently. 62,67–72 Here, we adopt a conventional division of 3 main facets of environmental enrichment, namely social interactions, structural complexity and physical exercise.1,2 This division is attractive because it allows extrapolation of findings between human and rodent studies, thereby enhancing translation between clinical and preclinical reports73 (Figure 1). We consider the interactions of each environmental component with the BBB and GB.
Social interactions
Humans are inherently social animals, and interaction with other individuals is a fundamental necessity, as much for emotional well-being as for normal development and physiology of the brain and neural circuits.56,74,75 Similarly, rodents participate in a complex array of social relationships that range from sexual interactions and family bonding to aggression and dominance hierarchies.76 Given that social encounters play such an important role in our lives, isolation is consequently linked to a host of unfavourable outcomes including risk of depression, cognitive decline, cardiovascular dysfunction, systemic inflammation and immune dysregulation. 56,74,77 Indeed, both social isolation and chronic social defeat stress disrupt neuronal reward circuits in the striatum of male and female mice.78,79 Evaluation of social interactions in humans is generally based on self-reported questionnaires, and tends to focus on subjective rather than objective measures of socialization. Far from being a shortfall of these studies, reports have suggested that physiologic effects are driven largely by perceived rather than actual social support or strain.77,80 In rodents, common manipulations include social stress through isolation or social defeat, while dominance in hierarchies is often assessed in pairs (for instance, with the tube test).38,81–83 Recently, advances in machine learning have enabled detailed tracking and analysis of maternal care and complex social interactions in animals.84,85
The physiologic outcomes of socialization extend far beyond the brain. In humans, for example, social support can buffer a stress-induced increase in concentrations of circulating glucocorticoids.86 Further, social interactions influence markers of systemic inflammation, with reported support from family being associated with a decrease in IL-6, while family strain increased serum levels of this cytokine. Support and strain from spousal relationships were associated with reduced or elevated levels of E-selectin, respectively.87 Interleukin-6 is a marker of systemic inflammation,38 and circulating E-selectin has been proposed as a marker of BBB permeability;49 thus, these findings suggest that positive social interactions can reduce immune response and promote barrier integrity. These fluctuations have also been observed in rodents; social instability in adolescent rats alters hippocampal gene expression and the composition of the gut microbiome, and social isolation in adult male CD-1 mice modifies the cytokine profile and behavioural response to immune challenge.88,89 Moreover, in social hierarchies among males, dominant CD-1 mice possess an immune profile and leukocyte transcriptional patterns that are distinct from submissive mice.90 These links between environmental, systemic and behavioural changes during social encounters imply communication of relevant information across the GB and BBB (Figure 3).

Mechanisms of environmental influence on the blood–brain and gut barriers. Social interaction, structural complexity and locomotor activity are distinct components of both the human and rodent environment that produce unique effects on the gut barrier, periphery and blood–brain barrier. AQP = aquaporin; Bdnf = brain-derived neurotrophic factor; bFgf = basic fibroblast growth factor; Cldn = claudin; CRH = corticotropin-releasing hormone; IL = interleukin; LPS = lipopolysaccharide; NF = nuclear factor; GSK = glycogen synthase kinase; HCAR = hydroxycarboxylic acid receptor; Ocln = occludin; SCFA = short-chain fatty acid; TNF = tumour necrosis factor; Vegf = vascular endothelial growth factor; ZO = zonula occludens.
Correspondingly, social experiences affect, and are affected by, sex-specific properties of cells in the neurovascular unit. For example, microglia, which are highly sensitive to environmental factors, monitor the generation of new cells during development and contribute to sex-specific social tendencies via pruning of newborn astrocytes in mice.91,92 Social isolation, meanwhile, appears to induce astrocyte reactivity, as evidenced by a report of elevated expression of glial fibrillary astrocytic protein in the hippocampus of isolated mice.93 A further study suggests that isolation impairs astrocytic connectivity, reducing cell body volume and process complexity.94 Although it has not been directly studied, socialization-related adaptations in these cells likely modify function of the neurovascular unit and BBB integrity to influence behavioural changes.95 In brain endothelial cells, models of social stress, including social isolation and social defeat stress, lead to dysregulation of tight junction proteins and subsequent loss of BBB integrity in both male and female mice.38,49,81,83,96 Interestingly, this effect is dependent on timescale; in rats, short duration of isolation stress (7 d) produces deficits in the tight junction protein claudin-5, while longer durations (6 wk) actually increase mRNA expression of claudin-5 in the male hippocampus versus the female prefrontal cortex.81,83 Furthermore, targeted downregulation of claudin-5 at the BBB is sufficient to produce deficits in social interaction in male and female C57BL/6 mice.38,49 Although these findings hint at a connection between BBB integrity and social activity, the intricacies of this relationship remain unclear. Recent reports are helping to develop a mechanistic insight; for instance, dominant and submissive CD-1 mice display distinct immunological profiles, with dominance associated with a shift toward the adaptive immune system and, therefore, an increase in B-and T-cell activity.90 Active T cells can cross the BBB and infiltrate the brain to communicate information, and it is possible that the BBB mediates communication between the brain and immune system to influence social status. It is also likely that BBB phenotype is important for sex-specific behaviours. An elegant study involving Drosophila suggests that signalling through the moody receptor (a melatonin receptor homologue) in the BBB is required for normal male courtship behaviour.97 In humans, melatonin possesses antioxidant qualities that are important for maintaining BBB integrity, and is also involved in sex-specific sexual development, although a definitive link between the 2 functions remains to be proven.98 Finally, chronic social defeat stress increases expression of histone deacetylase-1, reducing histone acetylation and, thus, inactivating transcription of genes, including claudin-5, which has been proposed as a mechanism for development of stress-induced deficits in social behaviours.99
The gut is also strongly implicated in social interactions. In humans, HPA activation by an acute social stressor like public speech leads to a transient permeabilization of the GB, as measured by differential urinary excretion of ingested, nonmetabolized sugars.100 This effect can also be elicited by injection of corticotropin-releasing hormone and, in both cases, is attenuated by an ingestible inhibitor of enteric mast cell degranulation. Meanwhile, perceived social exclusion alters the composition of the gut microbiome, favouring species of the genus Prevotella, which has been previously linked with emotion perception and limbic system connectivity.29,101,102 Interestingly, in a group of rats with colon cancer, social interactions, but not environmentally complex housing, was found to modulate gastrointestinal morphology by increasing expression of occludin, a tight junction protein, and IL-10, an anti-inflammatory cytokine, in the intestinal mucosa and blood plasma.103 These changes can influence the brain as seen in prairie voles, for which socialization produces lasting enrichment in diversity, which correlates with changes in region-specific brain transcription and glial cell abundance.104 On the other hand, social stress from long-term social isolation alters blood pressure and volume in the small intestine among female rats, along with expression of BBB-related tight junctions.83 Furthermore, mice susceptible to social defeat stress are characterized by changes in gene expression of proinflammatory cytokines IL1B and IL6 in the prefrontal cortex, in line with a higher abundance of Bacteroidetes, a genus related to gut dysbiosis. 105 A recent study suggests a mechanism for microbiome influence on social behaviour in which gut dysbiosis leads to diminished bacterial production of butyric acid — a prominent SCFA that normally diffuses across the GB and BBB to promote synaptic plasticity by inhibition of histone deacetylase 2 activity — thereby increasing transcriptional suppression and a subordinate state.106 These findings highlight that both the GB and BBB are important components in mediating communication between the brain, periphery and environment during social interactions.
Structural complexity
Structural enrichment is a paradigm stretching back to Donald O. Hebb and his pet rats.3 Novel and diverse environmental features offer a continual opportunity for informal learning and accelerate development of neural circuits and cerebral vascular networks.107,108 In humans, for example, high socioeconomic status is associated with better housing quality, leisure time and more opportunities for travel, leading to a greater frequency of novel experiences, compared with low socioeconomic status. Reproduction of this concept for preclinical studies with rodents varies widely between research groups, but generally involves larger caging, multiple levels in the environment, space for shelter, extra nesting material and manipulable objects or toys, which collectively provide an enhanced visual, olfactory and somatosensory landscape.2,66 Despite poor standardization and diversity of experimental setups, the behavioural and neural outcomes of environmental complexity in rodents have been remarkably consistent. Indeed, mice housed with some kind of structural enrichment generally show enhanced learning and memory performance,5 and diminished anxiety-like behaviour.109,110 One of the most common biological observations in rats and mice with complex housing is the genesis of new neurons in the hippocampus, 9 although it is unclear if this is sufficient to explain behavioural outcomes.2,111 Further neural consequences of this enrichment include increased dendritic branching, neuronal firing rate and synaptic plasticity in several cortical and striatal brain regions.6,7,9,112,113
However, it appears likely that systemic adaptations play an indispensable role in driving cognitive and behavioural effects of environmental complexity. Modifications to CD8+ T-cell transcription and inflammatory response, for example, appear to be essential components of environmental complexity–driven effects on hippocampal neurogenesis.16 Moreover, elevated levels of circulating SCFAs from shifts in microbiome composition are indispensable for positive effects of structural enrichment on neuronal plasticity and microglial function in the visual cortex.13 Finally, both novel and complex environmental features influence the responsiveness of the HPA axis. Enriched housing in CD-1 mice appears to slightly increase basal levels of circulating corticosterone; 114 furthermore, exposure to novel objects and larger cages elevates corticosterone in the blood of male C57BL/6 mice.115 The importance of these peripheral responses to the cognitive outcomes of structural enrichment suggests that some of this external information must cross the BBB and GB to reach the brain (Figure 3).
Blood vessels in the brain and the BBB are indeed shown to be actively modified by learning and memory tasks,115–118 indicating a role for the neurovasculature in mediating cognitive and behavioural outcomes of structural enrichment. Correspondingly, several reports have indicated that enhanced blood vessel size and density compensate for neuronal changes in rodents with complex housing.4,8,63,119 These cerebrovascular adaptations arise through an accumulation of minute effects of environmental complexity on various cells of the neurovascular unit. For example, complex housing increases astrocytic arborization and plasticity in the rat dentate gyrus,120 and promotes tighter astrocyte–synapse interactions, which could improve coupling between neurons and the neurovascular unit.121 Meanwhile, microglia from mice housed with structural enrichment exhibit diminished reactivity and potentially enhanced polarization toward a protective, anti-inflammatory phenotype.12,122 Accumulating evidence also highlights the role of growth factors in the effects of complex housing, such as brain-derived neurotrophic factor and fibroblast growth factor 2, both of which are known to influence endothelial proliferation and genesis of blood vessels.60,110,123–125 Further, preclinical studies that employ environmental complexity as a protective intervention in disease models implicate the BBB as an important site for preventive adaptations. Angiogenesis and blood vessel density have been linked to the protective effects of environmental complexity against several disease models in rodents, including stroke,126 diabetes127 and aging.128 In a set of experiments using the chronic cerebral hypoperfusion model of vascular dementia in rats, environmental complexity, combined with access to a running wheel, improved memory deficits and reliably prevented disease-related loss of tight junction proteins, namely claudin-5 and zonula occluden-1, suggesting a neuroprotective adaptation at the neurovascular unit.129–131 This protective effect appears to be mediated by inhibition of β-catenin phosphorylation132 and rescue of autophagy dysfunction.131
The GB also plays an important role in mediating the effects of complex environments. Indeed, alterations to the gut microbiome that are induced by complex housing combined with a running wheel facilitate environment-related synaptic plasticity and microglia arborization in the visual cortex via increased circulating levels of butyrate, an SCFA.13 Access to physical exercise, combined with a complex environment, increases microbial production of SCFAs, which appears to be a key factor in driving neurogenesis in the hippocampus, a feature observed with structural enrichment in mice.9 It also accounts for a portion of enrichment-related increases in expression of hippocampal growth factors.133 In disease conditions, mice in structurally enriched cages possess a distinct microbiome composition when compared with standard housing,134 which could represent a confounding factor in studies seeking mechanistic insights for brain disorders. A similar impact was observed for the immune system, with complex housing promoting an anti-inflammatory response in both the blood and gut mucosa.103 In fact, combining complex housing with a running wheel can enhance the intestinal epithelium’s response for wound repair in the context of colon cancer.135 Finally, environmental complexity can modulate gut microbiome composition by increasing levels of Lactobacillus, a bacterium associated with anti-inflammatory effects.136 These findings shed light on a possible role for the GB in mediating gut–brain communication in complex environments.
Physical exercise
Physical activity is one of the single most important modifiable factors for individual health and has been proposed as the critical environmental variable in regulating health outcomes. 71,72,137 Regular voluntary exercise is linked to several beneficial effects, including improvements to memory and learning, as well as decreased risk of cardiovascular and psychiatric diseases.57,138 Moreover, exercise is a strong natural reward that enhances neuronal activity in key brain areas for reward processing.139 However, despite the inherent motivation, environmental constraints profoundly influence the possibility for an organism to be active. In humans, for example, community levels of physical activity correlate positively with availability of parks and public spaces, and negatively with socioeconomic barriers to space for exercise. 140,141 Likewise, mice with free access to a running wheel will habitually run up to 15 km per day, a behaviour not observed in standard laboratory cages.142 Therefore, locomotion can be seen not just objectively as quantifiable movement, but more broadly as a measure of external influence on expressed behaviours, a shift that emphasizes the environmental aspects of physical exercise. Activity is commonly evaluated among human participants through questionnaires about lifestyle and exercise, or, in clinical studies, as an intervention, whereby a subset of patients performs a predetermined exercise regimen. Meanwhile, locomotion studies in rodents are limited to monitoring of regular movement within the home cage, or with access to a treadmill or running wheel.143
Although exercise is linked to many changes in the brain — including elevated striatal dopamine and enhanced synaptic plasticity and hippocampal neurogenesis — it also produces broad systemic effects.69,137,144 Among both humans and rodents, physical activity produces changes to the immune system, including a distinct cytokine profile that differs slightly but significantly from that elicited by infection or damage. In contrast to the pathogen response, strictly proinflammatory cytokines such as TNF-α and IL-1β are not significantly increased after physical exercise.138 Instead, IL-6 alone is heavily secreted from contracting skeletal muscles, promoting a downstream anti-inflammatory effect by stimulating release of IL-10.145,146 Further, exercise is linked to slight shifts in microbiome composition, generating a pronounced increase in circulating levels of butyrate.147,148 Glucocorticoid release is elicited by acute exercise in an intensity-dependent manner, and chronic exercise can elevate baseline levels of cortisol.149 Furthermore, a brief period of treadmill running dampens leukocyte sensitivity to elevated glucocorticoid levels and reduces risk of HPA over-reaction.150 However, forced physical exercise is detrimental, with similar outcomes as exposure to chronic stressors. It promotes higher expression of inflammatory markers and HPA reactivity, which in turn are related to anxiety-like behaviours.151 Paradoxically, voluntary exercise–induced elevation of IL-6 and corticosterone is also similar to the effects of chronic stress, but produces completely opposite effects on the brain.152 Although stress can be detrimental, voluntary exercise is often associated with beneficial outcomes including improved memory and reduced anxiety-like behaviours.64,69 An important difference that could explain the behavioural outcomes of physical exercise versus stress exposure, despite common biology, is controllability. In fact, controllable events might even promote control over unpredictable events153 and produce differential HPA activation or immune profiles.154,155 Interestingly, voluntary exercise can be seen as a controllable event that produces several changes to the BBB and GB (Figure 3). A better understanding of the impact of physical exercise on these barriers could help its promotion in the context of mental health or other conditions.
Among humans, physical activity produces an increase in both blood supply to the brain and blood oxygenation within minutes, especially in the prefrontal cortex and motor areas.156–158 In rodent models, long-lasting increases of blood flow caused by several weeks of physical exercise is associated with increased blood vessel density in the cortex, striatum and cerebellum.8,67,159,160 Exercise also strengthens interactions between cells of the neurovascular unit to produce a healthier BBB. For instance, access to treadmill exercise boosts astrocyte density in the hippocampus of male rats161 and in the prefrontal cortex of male C57BL/6 mice.162 Wheel running increases astrocyte reactivity, as measured by cell body area, and staining of aquaporin-4 — a water channel localized on astrocyte end-feet that is heavily involved in communication between astrocytes and endothelial cells of the BBB — in both the hippocampus and prefrontal cortex of male Wistar rats.163 Aquaporin-4 is involved in various functions such as regulation of extracellular space volume, cerebrospinal fluid circulation, waste clearance, neuroinflammation, calcium signalling, cell migration and more; thus, it is directly involved in BBB homeostasis and permeability.164 Physical exercise changes the cellular distribution of aquaporin-4, with higher concentration at the end-feet. 165 Increased astrocytic coverage of blood vessels was also reported in mice with access to physical exercise (30-min sessions, 3 times/wk for 12 wk).61 Furthermore, smooth muscle cells along cerebral capillaries, including pericytes, express high levels of a lactate receptor, hydrocarboxylic acid receptor-1, after long-term treadmill running among C57BL/6 mice. Subsequent downstream signalling of hydrocarboxylic acid receptor 1 leads to release of vascular endothelial growth factor (VEGF) and increased angiogenesis,160 a finding that coincides with earlier studies linking elevated VEGF expression to neurogenic and behavioural changes after exercise.166,167 This link provides evidence that the BBB acts as a mediator for cerebral and systemic effects of environmental change.
Physical exercise also promotes strength and maintenance of tight junctions between endothelial cells of the BBB and protects against damage-induced loss of barrier integrity. Among male C57BL/6 mice, access to a running wheel enhanced expression of occludin in the brain and prevented extravasation of injected tumour cells;168 among male Wistar rats, a graded treadmill exercise rescued expression of striatal claudin-5 after induction of type I diabetes.169 Souza and colleagues170 linked exercise-related protection of tight junction proteins to reductions in inflammatory signalling and oxidative stress of the central nervous system. The protective antioxidant effects of physical exercise in the brain appear to be mediated by inhibition of glycogen synthase kinase-3 (GSK-3), a redox-sensitive enzyme that destabilizes tight junction architecture via β-catenin phosphorylation.171 In vitro studies have found that inhibiting GSK-3 in brain endothelial cells reduces inflammatory signalling and improves the half-life of tight junction proteins, namely claudin-5 and occludin.172,173 In mice, voluntary wheel running could inhibit GSK-3 by increasing serum concentrations of the clusterin protein, which binds strongly to the low-density lipoprotein receptor-related protein 8 on microvascular endothelial cells in the brain.174 Activation of this receptor suppresses GSK-3 activity through a pathway known to promote BBB integrity in vitro.175,176
Excessive activity (> 60 min at > 70% of work capacity) can promote a leaky gut by activation of the HPA axis, along with elevated body temperature, leading to hypoperfusion of abdominal organs and subsequent changes in level of nutrients and immune signals reaching the GB.177,178 This effect produces a transient increase in permeability of the epithelial barrier, as measured with a sugar absorption test, as well as increased circulating markers of intestinal stress.178,179 In rodents, voluntary physical exercise influences inflammatory gene expression in intestinal leukocytes, downregulating proinflammatory cytokines while increasing levels of anti-inflammatory factors and antioxidant enzymes.180,181 Similarly, in humans, short periods of exercise increase lymphocyte levels in the blood, but diminish expression of toll-like receptors, suggesting that physical activity can modulate the immune response to pathogens and stressors.182,183 Physical exercise increases diversity of the gut microbiome, according to a direct comparison of samples from athletes and people with a sedentary lifestyle.184 This effect may be mediated by innate immune cells that condition the gut microbiome toward optimal composition of serotonin and SCFA-producing bacteria.147,185 The microbiome of people with low body weight despite a sedentary lifestyle is similar to that elicited by exercise, suggesting that a specific bacterial profile could be associated with physical health.186 Indeed, randomized controlled trials involving children with obesity showed that deleterious gut microbiome and proinflammatory signalling could be reversed to the levels of healthy children via a physical exercise intervention.187 As for adults with obesity, 6 weeks of physical exercise was sufficient to modify the diversity and composition of the gut microbiome by increasing SCFAs and butyrate-producing bacteria in relation to body mass and loss of weight.188 At the mechanistic level, butyrate was found to be responsible for assembly and maintenance of tight junction proteins in cultured human intestinal epithelial cells,189 suggesting that exercise-induced microbial production of butyrate could promote a healthy GB.
Barrier–environment interactions in development and aging
Most of this evidence describes how environmental components influence the normal functioning of the BBB and GB during adult life; however, they do not explicitly consider the impact of early challenges or the long-term effects of environmental conditions on barrier function later in life. Across the lifespan, from the maternal environment to life as an older adult, environmental conditions have the potential to modify barrier functioning; indeed, enriched environments may improve function and the expression of anti-inflammatory factors, whereas detrimental environmental conditions could lead to improper functioning and exacerbated inflammation. 73,190,191 Brain maturation and development is a continuous process; different developmental stages have characteristic behavioural, neurologic and immune profiles. This progression between life stages is similar across humans and rodents and is mainly mediated by preprogrammed physiologic changes in cell and hormonal activity.192 Throughout development, there are periods (e.g., in embryo, childhood, adolescence) of increased sensitivity to the surrounding environment’s threats, like stress,40 as well as to beneficial, stimulating conditions. Traditionally, these windows of sensitivity are thought to be mainly determined by synaptic changes or neuronal growth, but the BBB and the GB also have specific developmental milestones during those sensitive periods. For instance, during the first postnatal days there is an increase in both astrocyte expression in the brain and microbial colonization of the gut that will help sustain normal neuronal wiring and cognitive function.30,33 Here, we discuss how the function of the barriers can be improved or harmed by environmental features from a lifespan and life stage perspective (further details about BBB development can be found elsewhere)30,33,193,194. In addition, we highlight research opportunities for which a developmental perspective may contribute to understanding of the relationship between barrier function and mental health.
Embryogenesis and development
During the first embryonic or fetal stages (embryonic day 10 in rodents and gestational week 8 in humans) the permeability of the BBB is set to protect the growing brain against potential threats,195 and the GB prepares for complex diets by triggering specific cell differentiation of the intestine.194 However, the maternal environment, in combination with embryonal or fetal age, can modify the development of these barriers and their susceptibility to factors such as toxins,196,197 viruses198,199 or unhealthy diet.200 Environmental modification in the sociability, complexity (e.g., socioeconomic status in humans) and physical activity of the mother during pregnancy can produce positive or negative transgenerational effects by setting a specific proinflammatory footprint in the offspring.201–204 These modifications may even contribute to future stress reactivity by changing release of stress hormones and expression of brain glucocorticoid receptors.205,206 For instance, environmental enrichment by structural complexity during pregnancy alters inflammatory responses mediated by circulating cytokines (IL-6, IL-10 and TNF-α) and lipopolysaccharide-induced expression of IL-22 and prostaglandin E2.207 This immune response profile promotes healthy offspring and may help with the normal development of motor skills. Likewise, in a transgenic mouse model of Alzheimer disease, physical exercise during pregnancy showed long-term effects on the offspring’s neurovascular function by improving cerebral oxygen and nutrient supply, increasing angiogenesis and BBB transporter activity and reducing microglia reactivity.208 Furthermore, positive environmental conditions for the mother can reduce behavioural outcomes associated with anxiety during the offspring’s adolescence or adult life.209 On the other hand, a detrimental environment such as social isolation and reduced complexity during pregnancy may negate the beneficial effects on synaptic transmission driven by access to an enrichment environment. 210 Housing mothers with a whole ensemble of enrichment during pregnancy — combining social interaction, physical exercise and structural complexity — can prevent the metabolic disturbances generated by hypoxic–ischemic events in the offspring’s early life and their impact in adult memory impairment.211 Positive cognitive outcomes are associated with long-term increases in the expression of growth-related proteins, downregulation of amyloid precursors and proper glucose metabolism. Considering the crucial role of BBB glucose transporters to meet the brain’s high energy demands, environmental enrichment may promote BBB-related transgenerational features that support brain metabolism and, ultimately, plasticity.
After birth, the growing BBB and GB undergo additional critical periods before they reach full maturation and functioning. Environmental modifications during postnatal and early-life periods — childhood and adolescence — play key roles in improving or altering barriers’ properties in the short and long term. They have been mostly investigated using paradigms of early-life adversity, such as maternal separation, limited besting and nesting, social isolation or sensorial deprivation, and as short- and long-term effects centred on brain connectivity and neuronal function.51,212,213 However, these postnatal periods also see expansion and refinement of the brain vascular architecture, with changes in endothelial transport and astrocyte expression.30,214 These features of BBB development suggest that neurovascular function may be sensitive to positive and negative environmental interventions; thus, access to an enriched environment early in life could potentially exert lasting changes in brain and behaviour. Indeed, structural complexity during postnatal development is associated with increased vascular branching and expression of the angiogenic VEGF and glucose transporters, at least in the rodent visual cortex.107,215 Structural complexity with VEGF administration, either in the visual cortex or in the hippocampus dentate gyrus, improves execution time and performance in a spatial memory task.117 Intriguingly, opportunities for social interactions after stressful events during adolescence counteracts deleterious stress effects by reducing aggressivity and anxiety- and depression-like behaviours in adulthood. These behavioural adaptations appear to be related to changes in corticosterone levels and increased expression of cell adhesion molecules associated with cell migration and outgrowth during development.216 Maternal separation can induce visceral hypersensitivity in rats, which can be improved via nutritional approaches early in life;217 however, how access to an enriched environment, physical exercise or social interactions could affect GB development remain understudied.
Later life and aging
On the other end of the lifespan, aging is generally associated with progressive decline of cognitive and physical health capacities, with cellular and molecular changes driving metabolic impairment, synaptic loss or alterations, neuroendocrine senescence and cerebrovascular decline.31,218,219 Onset of aging-related pathologies is linked with complex interactions between genetic and environmental factors, which can alleviate or exacerbate programmed senescence process for various systems or cell types that might increase susceptibility for conditions like Alzheimer or Parkinson diseases.31,219 Core features of age-related cognitive impairments, including dementias, are vascular dysfunction and BBB hyperpermeability through loss of occludin, zonula occluden-1, astrocytes and pericytes.220,221 Despite the progressive decline of cognitive processes, access to environmental enrichment during aging has positive effects in delaying onset of several pathologies, sustaining neurovascular integrity and activity, and preserving function of other brain systems during normal aging.62,73,222 Complementary environmental features can counteract age-related impairments. For instance, the whole ensemble of social, structural and locomotor enrichment increases lifespan and reduces anxiety-like behaviour, which appear to be associated with proper neurovascular function as measured by high glucose metabolism, expression of angiogenic markers and reduced expression of inflammatory genes in the brain.119,223–225 Interestingly, voluntary physical exercise during middle or older age promotes expression of the proangiogenic growth factor, VEGF; prevents age-related astrocyte, pericyte and myelin loss in the hippocampus; and reduces vascular leakage, possibly through higher vascular coverage by pericytes. 223,225 These beneficial effects of voluntary physical exercise on neurovascular health are associated with conserved motivation to engage in spontaneous behaviours, an indirect measure of cognitive function.225 Furthermore, long periods of social interaction during the transition from middle age to older age improves rodent performance in the novel object recognition task, which appears to be driven by alterations in corticosterone levels and reduced accumulation of vascular amyloid depositions.62 These findings support an extended window of opportunity during aging to intervene with the goal of sustaining BBB and GB integrity and promoting optimal function, leading to proper mental and physical health during this life stage.
Environmental enrichment can act as a preventive strategy to overcome the negative short- and long-term effects of stressful early-life events and age-programmed senescence on stress response, cell loss and decline of neurovascular function.53,73,226 Its positive outcomes seem to be mediated by changes in glucose metabolism, angiogenic factors that control vascular integrity and glial cell reactivity, leading to reduce maladaptive behaviours and preservation of cognitive function throughout the lifespan.117,191,227,228 It seems that vascularization and BBB integrity are complementary factors relevant to the promotion of learning and memory processes or emotional regulation.117 Most current research in this area has focused on neurodevelopmental disorders such as neonatal hypoxia or neurodegenerative conditions such as Alzheimer disease or dementia. Moreover, these studies have involved only a small subset of brain areas, highlighting the need to expand knowledge about how positive environment influences development and describe its role for other behavioural domains, such as mood disorders. Another research avenue will be to determine the role of each component of environmental enrichment in development, as most of the research to date has focused on the general effect of the enrichment environment without considering the effects of each feature independently or comparing them with each other. Few studies have directly compared beneficial effects of one environmental feature with other features or the summative effect of all components; thus, it is impossible to rule out that single strategies may be more effective in specific contexts.62 Identification of the main element leading to behavioural improvement will help to establish translational low-cost, highly effective strategies in the promotion of healthy lifestyle and improved mental health.
Barrier–environment interactions in mood disorder pathogenesis
The importance of studying environment–barrier interactions is heightened in the context of brain disorders. Psychiatric conditions like MDD, bipolar disorder and schizophrenia rank among the leading causes of disability worldwide, affecting nearly 1 in 4 people.229 Despite high prevalences — 7% for MDD, 3% for bipolar disorder and 1% for schizophrenia — and severe effects on quality of life, commercially available therapeutics have advanced only slightly in the past 30 years, while between 30%–50% of all affected individuals are slightly responsive or nonresponsive to current treatments. 65,230 Environment constitutes an important proportion of risk for these diseases, with genetic contributions estimated at 60% of risk for schizophrenia to as low as 30% for MDD.231 This suggests disruption of pathways for environment–brain communication, like the BBB and GB, in the pathogenesis of mood disorders. Correspondingly, loss of barrier integrity is increasingly recognized as a hallmark feature of neurodegenerative disorders, with dysfunction of BBB and GB reported among people with MDD,36,38,49,232,233 bipolar disorder45,46 and schizophrenia.44,47,234 Despite the well-known involvement of the environment, BBB and GB in the pathogenesis of mood disorders, the role of communication between these systems remains unclear. This section discusses a possible role for environment–barrier interactions in progression of psychiatric conditions and identifies areas for future research.
Environment as a source of stress
Past and present environments can promote or prevent the onset of mental health disorders through modification of the stress response. Negative or impoverished conditions — such as low socioeconomic status, social isolation and sedentary lifestyle — are sources of chronic stress linked to peripheral inflammation, 55,235 gut dysbiosis236 and risk for development of mood disorders.54,56 However, positive and stimulating environmental conditions also confer a degree of stress; social interactions, structural complexity and physical exercise all increase HPA activation (Figure 3), and yet are associated with beneficial effects on learning and memory via adaptations in the BBB and GB. Moreover, preclinical studies have found anti-depressant effects of physical exercise and complex housing in the context of social defeat stress.58,59 What determines whether the influence of a given environment is positive or negative?
Part of the answer comes from the inherent properties of the stress response, which is described as an allostatic system, meaning it can adapt to environmental demands, changing its equilibrium point through sustained alterations in allostatic mediators like cytokines and glucocorticoids.237 The dynamic nature of allostatic systems is crucial to maintaining homeostatic mechanisms during periods of external change.238,239 However, in the presence of a severe or prolonged stressor, these systems are susceptible to allostatic overload, at which point the capacity to maintain stability is exceeded, resulting in illness or death.237,240 Importantly, in the face of a given stress, some people will experience allostatic overload and pathology, while others will be able to cope and return to baseline. This capacity for returning to baseline after stress is termed resilience, a concept that has come to shape the way health and disease are conceptualized.241,242 In the case of psychiatric illness, the stress response is elevated to the point of damaging the BBB to induce neuroinflammation and cognitive deficits,38 while a subthreshold, or “good,” stress, can increase the functional range of the system and enable an organism to weather greater fluctuations in the future.1 This framework clearly explains how a stimulating environment could act as a low-grade stressor to actually enhance an organism’s capacity to deal with future challenges — a phenomenon that has been referred to both as buffering56 and inoculating against1 the effects of chronic stress.
Barrier–environment interactions mediate stress resilience
As critical frontiers for communication between the environment, periphery and brain, the BBB and GB are key loci for adaptations that determine the influence of the environment on cognition and behaviour. These barriers are known to play a crucial role in mediating resilience versus susceptibility to chronic stress. For instance, inflammation-related epigenetic suppression of the claudin-5 promoter in endothelial cells of the BBB is a key predictor of susceptibility to social defeat stress in male mice.99 Among female mice, 6 days of chronic variable stress alters gut microbiome composition and disrupts GB integrity, resulting in elevated levels of circulating llipopolysaccharide-binding protein and behavioural deficits.25,36,49 However, the involvement of the BBB and GB in effects of positive environments on stress response and mental health is less clear. One preclinical study reported that peripheral injection of fibroblast growth factor-2, a growth factor associated with BBB integrity and positive cognitive outcomes of structural enrichment, is sufficient to diminish anxious behaviour in a study of selectively bred, high-anxiety rats.110 In another report, physical exercise prevented depression-like behaviour that would normally have been induced by chronic unpredictable stress exposure by increasing astrocyte density and proliferation in the hippocampus. 243 Proliferation of astrocytes can be associated with more extensive astrocyte covering of blood vessels, a key feature of BBB integrity.20 This finding corresponds with results from a rat model of vascular dementia in which physical exercise prevented cognitive deficits by increasing astrocyte coverage of blood vessels.61 Further, in mice, oral administration of SCFAs — which are produced in higher levels by the gut after structural enrichment, social interactions and physical exercise — promoted anxiolytic-like behaviours and prevented social stress–inducing anhedonia, stress reactivity and increased intestinal permeability.244 Interestingly, it is known that SCFAs exert their effect mainly through free fatty acid receptor-2 and −3; however, studies of their role in mediating the gut–brain axis have focused on receptor modulation with chemogenetic tools or the development of allosteric modulators without behavioural evaluation;245 thus, it will be intriguing to evaluate the impact of these SCFA-related chemogenetic tools and allosteric modulators in the context of stress-related disorders. In humans, the finding that social support is associated with reductions in circulating levels of E-selectin is striking,87 as this molecule has been reported as a sex-specific biomarker for BBB disruption associated with MDD.49 Moreover, reports have linked socioeconomic status with gut and microbiome health,236 as well as better measurements of intestinal barrier integrity via a sugar permeability assay in children.246 Although evidence is scarce, it is increasingly apparent that environmental modifications of the neurovasculature and gut epithelium are important in development of stress resilience.
This highlights a new opportunity for drug discovery, as the BBB and GB are increasingly seen as potential targets for novel antidepressant and antipsychotic therapies. Drug discovery programs that aim to reduce stress-induced damage by targeting pathological features of mood disorders are an obvious approach. For example, 2 recent preclinical studies — one employing pharmacological modulation of Wnt-7a signalling with the Gpr124/Reck agonist in stroke models, and another regulating the transforming growth factor-β pathway in experimental epilepsy using RepSox, an inhibitor of activin receptor-like kinase-5 — have shown that BBB integrity can be recovered by indirect upregulation of claudin-5 in brain blood vessels.247,248 However, these target-driven studies are limited in scope, focusing strictly on repair and thereby missing out on bigger-picture strategies associated with overall mental well-being.249 The proresilient effects of social interaction, environmental complexity and physical activity offer an alternative, phenotype-based method for drug development, making it possible to investigate pathways associated with prevention instead of treatment or to investigate stress-related changes underlying cognitive deficits and mood disorders. Several potential antidepressant targets can be gleaned from environment–barrier interactions (Figure 2), including levels of circulating factors like cytokines and SCFAs, expression of growth factors in the brain and activity of redox-sensitive enzymes like GSK-3 in brain endothelial cells. For decades, GSK-3 has been targeted in the context of psychiatric conditions, notably with lithium.250 The efficiency of antidepressant treatment has been linked to an inhibitory effect on GSK-3 activity; 251 nevertheless, its relevance in the treatment of MDD is unclear. Greater attention should be paid to these pathways in future studies to elucidate potential roles in promoting stress resilience at the BBB and GB. Ultimately, a better understanding of environment–barrier interactions in the context of psychiatric disorders will greatly improve understanding of these diseases, as well as capacity to prevent and treat them.
Conclusion
In the early 20th century, it was commonly held that the adult brain was a rigid organ, independent of external influence (Figure 4). The experiments of Donald O. Hebb3 helped to strike down this dogma and repopularize the idea that environment can change the brain, a concept that reaches back centuries.5,252 His findings of environmental influence on learning and memory in rats lent enormous support to the theory of synaptic plasticity, which ultimately changed the scope of future neuroscience; yet, this revolution was narrow-sighted, and the focus on neurons and the brain left systemic aspects of enrichment sidelined for years. Alhough it has been long known that environment can influence the immune system, circulating glucocorticoids and microbiome composition,14,52,146,148,253 only recently have the interactions between these systems and the brain been investigated. 13,16,88,114 As critical mediators of communication between anatomic compartments, biological barriers like the BBB and GB are major interfaces for translating environmental information between the brain and periphery.
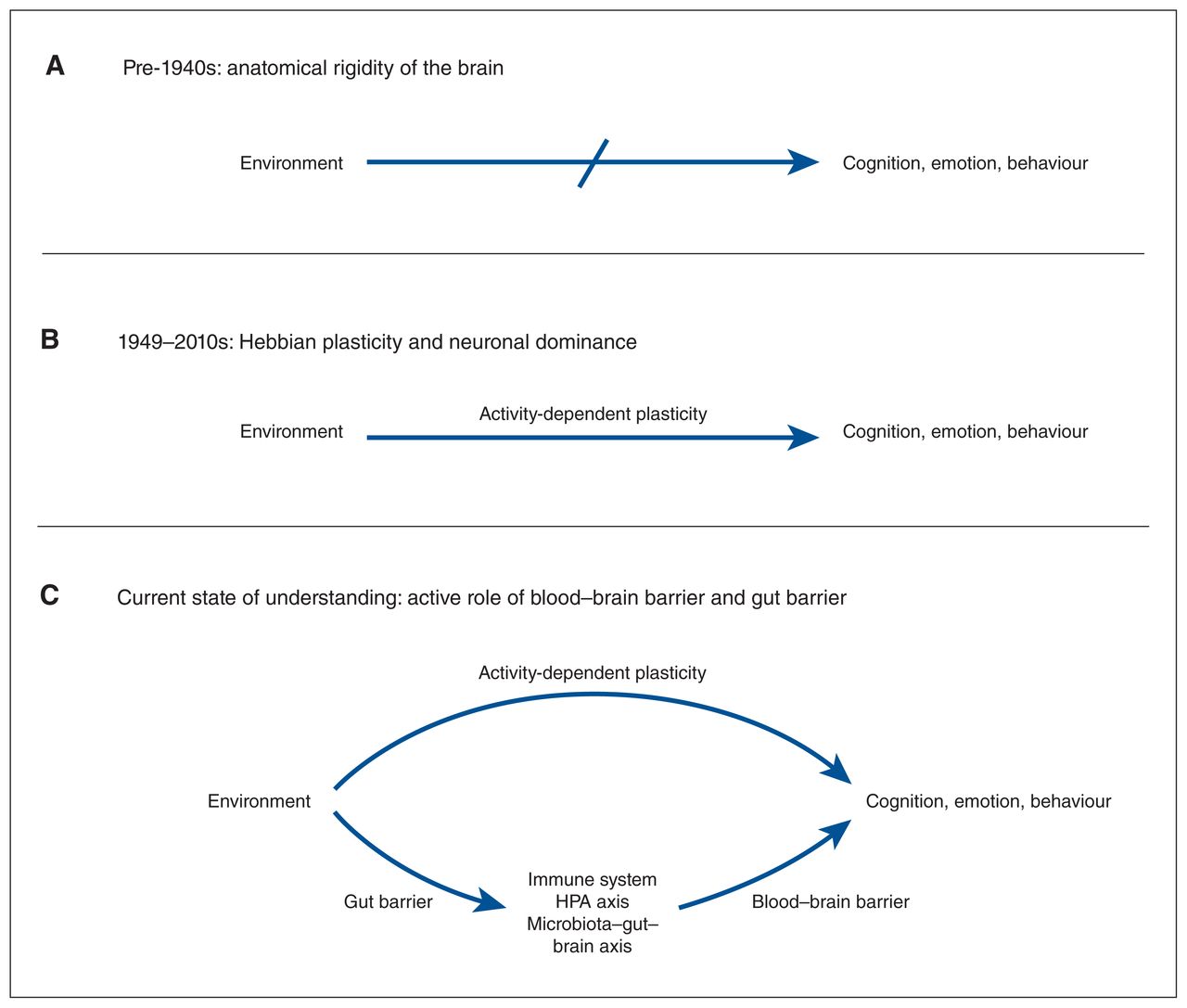
Historical development of understanding of environmental influence on cognition, emotion and behaviour. (A) For the first half of the 20th century, it was widely believed that the adult brain was a rigid organ, insensitive to change from external factors. (B) Afterr Hebb’s demonstration of positive effects of a stimulating environment on learning and memory, it was established that environmental influence on neuronal plasticity and firing was responsible for long-term changes in mental state. (C) In recent years, systemic outcomes have been increasingly recognized as indispensable for positive outcomes of environmental enrichment. Thus, peripheral signals, mediated by the blood–brain and gut barriers, are important features acting in parallel to central plasticity changes to influence cognition, emotion and behaviour in response to shifting environments. HPA = hypothalamic–pituitary–adrenal.
In this review, we have summarized a broad literature that describes multifaceted interactions between the environment, the BBB and the GB to argue that these barriers play a central, active role in external modulation of cognition, emotion and behaviour throughout the lifespan. Further, we emphasized an important early role for environment–barrier interactions in the pathogenesis of mood disorders and suggested potential targets for novel therapeutics. We focused our discussion on 3 major environmental components that have been identified as the key loci for environmental influence on cognition, behaviour and underlying neurobiology — social interactions, structural complexity and locomotion.1,2,62,68,70 This focus meant that several other factors such as environmental pollution, diet, temperature, auditory stimulation and drug and alcohol consumption — all of which are also known to affect behaviour and cognition — were beyond the scope of this review. However, these are also important variables that can modify barrier integrity and should not be neglected by future studies. In addition, we found that the literature lacked a standardized paradigm for studying environment in a laboratory setting, such as cage components used as structural enrichment, and duration of exposure to running wheels or treadmill exercise. This variability impedes comparison of results; moving forward, we suggest clear and transparent reporting of environmental conditions, as well as adoption of standardized paradigms, which will be crucial to future detailed investigations of environment–barrier interactions in health and disease.
In a world where the number of diagnoses for psychiatric illnesses is growing in parallel with accumulation of environmental pollutants, feelings of loneliness and isolation, vast cultural and dietary shifts, sprawling suburbanization and more frequent extreme weather events that drive mass displacement, understanding the detailed interactions between environment and the brain should be of utmost importance, The BBB and GB are key loci for these investigations. Several questions remain unanswered, representing exciting opportunities for new research. We suggest 2 main areas of interest, namely characterizing relationships between socioeconomic status, social interactions and physical activity with BBB and GB health in human psychiatric disorders, and investigating molecular pathways involved in environment-associated protective adaptations at the BBB and gut epithelium for novel pharmaceutical targets. A fuller understanding of environment–barrier interactions will help in the development of effective therapies and preventive strategies for psychiatric conditions, as well as a better appreciation of how humans are affected by the worlds they inhabit.
Footnotes
↵* Share first authorship.
Competing interests: None declared.
Contributors: All of the authors contributed to the conception and design of the work, drafted the manuscript, revised it critically for important intellectual content, gave final approval of the version to be published and agreed to be accountable for all aspects of the work.
Funding: This work was supported by the Canadian Institutes for Health Research (Project Grant No. 427011 to Caroline Menard), Fonds de recherche du Québec — Santé (Junior 2 salary award to Caroline Menard), Brain Canada 2019 Future Leaders in Canadian Brain Research (Caroline Menard) and Canada First Research Excellence Fund (Sentinel North Research Chair to Caroline Menard). Sam Paton and José Solano are funded by Canadian Institutes for Health Research MSc and NeuroQuebec PhD scholarships, respectively.
- Received November 30, 2022.
- Revision received March 1, 2023.
- Accepted March 19, 2023.
This is an Open Access article distributed in accordance with the terms of the Creative Commons Attribution (CC BY-NC-ND 4.0) licence, which permits use, distribution and reproduction in any medium, provided that the original publication is properly cited, the use is noncommercial (i.e., research or educational use), and no modifications or adaptations are made. See: https://creativecommons.org/licenses/by-nc-nd/4.0/
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Article tools


