

Abstract
Background: The only antidepressant drugs that are effective in the treatment of obsessive–compulsive disorder (OCD) are those that effectively block the reuptake of serotonin (5-hydroxytryptamine; 5-HT). In humans, positron emission tomography studies have implicated the orbitofrontal cortex (OFC) in the mediation of OCD symptoms. In animals, administration of selective serotonin reuptake inhibitors (SSRIs) for 8 weeks (but not 3 weeks) led to increased release of 5-HT in the OFC, because of desensitization of the terminal 5-HT autoreceptors. However, the increase in synaptic levels of 5-HT in the OFC after long-term administration of SSRIs might be cancelled out by desensitization of postsynaptic 5-HT receptors. This study was undertaken to investigate if these OFC receptors adapt under such conditions.
Methods: In vivo electrophysiologic techniques were used in this animal study. Male Sprague–Dawley rats received the SSRI paroxetine or vehicle control, delivered by implanted osmotic minipumps, for 3 or 8 weeks. With the rats under anesthesia, neuronal responsiveness to the microiontophoretic application of various drugs was assessed by determining the number of spikes suppressed per nanoampere of ejection current.
Results: After administration of paroxetine for either 3 weeks or 8 weeks, there was no modification in the inhibitory effect of 5-HT, the preferential 5-HT2A receptor agonist (+)-1-(4-iodo-2,5-dimethoxyphenyl)-2-aminopropane hydrochloride (DOI) or the preferential 5-HT2C receptor agonist 3-chlorophenyl piperazine dihydrochloride (mCPP). In contrast, the inhibitory effect of the 5-HT1A receptor agonist 8-hydroxy-2-(di- n-propilamino)-tetralin (8-OH-DPAT) was attenuated in the OFC after both 3 and 8 weeks of paroxetine administration.
Conclusion: These results indicate a desensitization of postsynaptic 5-HT1A receptors in the OFC but a lack of compensatory adaptation of the 5-HT receptor(s) mediating the main effect of 5-HT in this brain region. These observations imply that the activation of normosensitive postsynaptic 5-HT2-like receptors may mediate the effect of enhanced 5-HT release in the OFC.
Introduction
The predominant neurobiologic hypothesis regarding the pathophysiology of obsessive–compulsive disorder (OCD) is that at least some aspects of the disorder are related to abnormal regulation of brain serotonergic function.1 However, this hypothesis was derived mainly from therapeutic studies demonstrating the efficacy of the tricyclic antidepressant and potent serotonin (5-hydroxytryptamine; 5-HT) reuptake inhibitor clomipramine in the treatment of OCD.2,3 These initial observations have been supported by subsequent investigations showing that other drugs with a potent inhibitory effect on 5-HT reuptake not belonging to the tricyclic family, such as paroxetine and fluoxetine, are effective antiobsessional agents, but drugs without potent 5-HT reuptake properties, such as desipramine, are not.4–6 On the one hand, clinical studies have shown that treatment with the 5-HT reuptake inhibitors clomipramine and fluoxetine leads to a reduction of activity in human prefrontal cortex areas, particularly the orbitofrontal cortex (OFC)3,7 and the head of caudate nucleus.8,9 Provocative stimuli inducing OCD symptoms increase regional cerebral blood flow in the OFC and the head of caudate nucleus. It is noteworthy that the reduction of OFC and caudate nucleus activity has been associated with clinical improvement in patients with OCD who responded to either pharmacotherapy or psychotherapy.9,10 On the other hand, preclinical investigations designed to assess 5-HT function have found that guinea pigs treated with selective serotonin reuptake inhibitors (SSRIs) have increased 5-HT release occurring through terminal 5-HT1D autoreceptor desensitization in the OFC but not in the caudate nucleus. In addition, it has been shown that 5-HT release is enhanced in the OFC after an 8-week but not a 3-week treatment with a different SSRI, such as paroxetine or fluoxetine, but not by the monoamine oxidase type A inhibitor moclobemide or electroconvulsive shocks, the latter 2 of which are ineffective in OCD.11,12
It is important to emphasize not only that alterations of presynaptic neurotransmitter components can underlie the effectiveness of drug treatment but also that the responsiveness of postsynaptic 5-HT receptors may contribute to, as well as altering, overall 5-HT transmission. Various classes of antidepressant treatments enhance 5-HT neurotransmission in the rat hippocampus with a time course that is consistent with their delayed therapeutic effect. This action would be mediated by different mechanisms such as postsynaptic sensitization to 5-HT, desensitization of the somatodendritic or terminal 5-HT autoreceptors, or both, or a desensitization of α2-adrenergic heteroreceptors located on 5-HT terminals.13,14 Long-term administration of SSRIs results in the desensitization of somatodendritic 5-HT1A autoreceptor function in the dorsal raphe, thereby allowing the firing rate of the 5-HT neurons to recover in the presence of the drugs, which in turn leads to an increase in 5-HT neurotransmission in the hippocampus.13,15 Although similar regimens result in the desensitization of physiologic responses mediated by postsynaptic 5-HT1A receptors in some brain regions such as the hypothalamus,16–18 electrophysiologic studies indicate that the sensitivity of postsynaptic 5-HT1A receptor-mediated responses in the hippocampus is not changed by SSRIs.13
The present study aimed to assess the sensitivity of postsynaptic 5-HT receptors in the OFC after 3- and 8-week treatments with paroxetine. The OFC is part of the circuitry implicated in OCD, and it is already known that 5-HT release in this structure is increased after 8 weeks of SSRI administration. The responsiveness to 5-HT itself was examined, as well as the responsiveness to the 5-HT1A receptor agonist 8-hydroxy-2-(di-n-propilamino)-tetralin (8-OH-DPAT), the preferential 5-HT2A receptor agonist (+)-1-(4-iodo-2,5-dimethoxyphenyl)-2-aminopropane hydrochloride (DOI) and the preferential 5-HT2C receptor agonist 3-chlorophenyl piperazine dihydrochloride (mCPP).
Materials and methods
Animals
Male Sprague–Dawley rats, weighing 250–350 g, were obtained from Charles River (St. Constant, Que.) and were kept on a 12:12 hour light–dark cycle under controlled conditions with regular indoor temperature and humidity. The animals had free access to standard rodent food (Charles River) and water during the study.
Treatments
The rats were anesthetized with halothane, and osmotic minipumps (Alza, Palo Alto, Calif.) were implanted subcutaneously; the minipumps delivered paroxetine (10 mg/kg per day) or the vehicle used to dilute this drug (50:50 mixture of ethanol and water) (in the controls). For the 8-week treatment, a new minipump was installed 4 weeks after the first implantation. The electrophysiology experiments were performed with the minipumps in place. For each treated rat, a control rat from the same batch was also studied on the same day, to prevent any variations in the effects of the drugs. All experiments were performed with the approval of the Uni versity Animal Care Committee and complied with rules set forth in the Guide to the Care and Use of Experimental Animals (1993) of the Canadian Council on Animal Care.
Single-unit recording and microiontophoresis
The rats were anesthetized with chloral hydrate (400 mg/kg i.p., followed by doses of 100 mg/kg as necessary to maintain complete anesthesia) and placed in a stereotaxic frame. Body temperature was maintained at 36°C–37°C with a thermostatically controlled heating pad. Extracellular unitary activity was amplified and displayed on an oscilloscope. Action potentials were discriminated from background noise by means of a differential amplitude discriminator, and the frequency of discharge was integrated over 10-second intervals with a ratemeter. Five-barrel micropipettes used for microiontophoresis were pulled with a vertical electrode puller and were broken back under a microscope to a diameter of approximately 5–9 μm. The stereotaxic coordinates for the OFC were as follows: A, 3.7 mm; L, 2.0 mm from the bregma; H, 2.5–4 mm below the cortical surface.19 The coordinates for the OFC in rats correspond to those for humans, which delineate Brodman area 47.20 At the end of the experiment, the final recording site was marked by passing a cathodal current through the recording barrel to deposit fast green dye such that the recording sites could be confirmed histologically. Because most OFC neurons are not spontaneously active under chloral hydrate anesthesia, a leak or a small ejection current of quisqualate (QUIS) was used to activate them within their physiologic firing range.21 The central barrel contained 2 mmol/L NaCl for recording, as did one of the side barrels for current balancing. Another side barrel contained QUIS (1.5 mmol/L in 400 mmol/L NaCl, pH 8; Tocris Cookson, Ballwin, Mo.). The remaining barrels were filled with 2 of the following drugs, all in 200 mmol/L NaCl adjusted to pH 4.0: 5-hydroxytryptamine creatinine sulfate (5-HT, 20 mmol/L; Sigma, St. Louis, Mo.), 8-OH-DPAT (20 mmol/L; RBI, Natick, Mass.), DOI (50 mmol/L; RBI, Natick, Mass.), mCPP (20 mmol/L; Bristol-Myers Squibb, Wallingford, Conn.). Neuronal responsiveness to the microiontophoretic application of various drugs was assessed by determining the number of spikes suppressed per nanoampere of ejection current.
Statistics
All results are expressed as mean (and standard error of the mean [SEM]). The magnitude of inhibition of different agonists was compared between control and treated rats with the 2-tailed Student’s t test (nonpaired). Differences were considered significant at p < 0.05.
Results
OFC discharge rates were not significantly different in control and treated rats (control rats, 10.3 [SEM 0.5] spikes/s, n = 104; treated rats, 11.1 [SEM 0.8] spikes/s, n = 112; p > 0.3), and no significant alteration in the ejection current of QUIS, used to activate these neurons, was observed in any group of rats (control rats, −80 [SEM 11] nA, treated rats, −70 [SEM 15] nA; p > 0.5).
It has previously been demonstrated that the in vivo microiontophoretic application of 5-HT, DOI and mCPP onto OFC neurons produces a suppressant effect on firing activity in rats,21 guinea pigs22 and mice.23 In the present study, all OFC neurons tested with 5-HT, 8-OH-DPAT, DOI and mCPP induced a reduction of firing activity (Fig. 1 and Fig. 2). This suppressant effect occurred in the absence of any alteration in the shape of the action potentials.

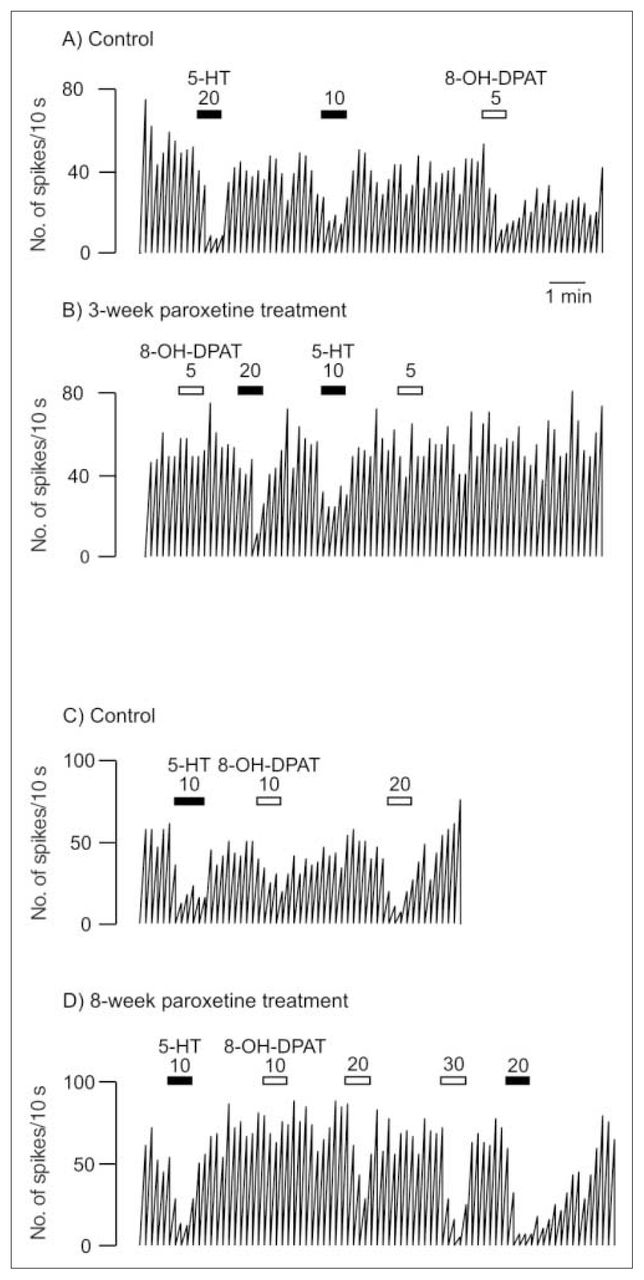
Integrated firing rate histogram of an orbitofrontal cortex neuron, showing its responsiveness to microiontophoretic application of serotonin (5-HT) and 8-OH-DPAT in control animals and rats treated with paroxetine for 3 and 8 weeks. Horizontal bars indicate the duration of the applications (currents given in nanoamperes). The time base in A applies to all traces. Traces in the control and treated rats were obtained from animals from the same shipment that were treated simultaneously after administration of vehicle or paroxetine. These traces were all obtained on the same day. Note the decreased responses to 8-OH-DPAT and the unaltered response to 5-HT in the treated rats.
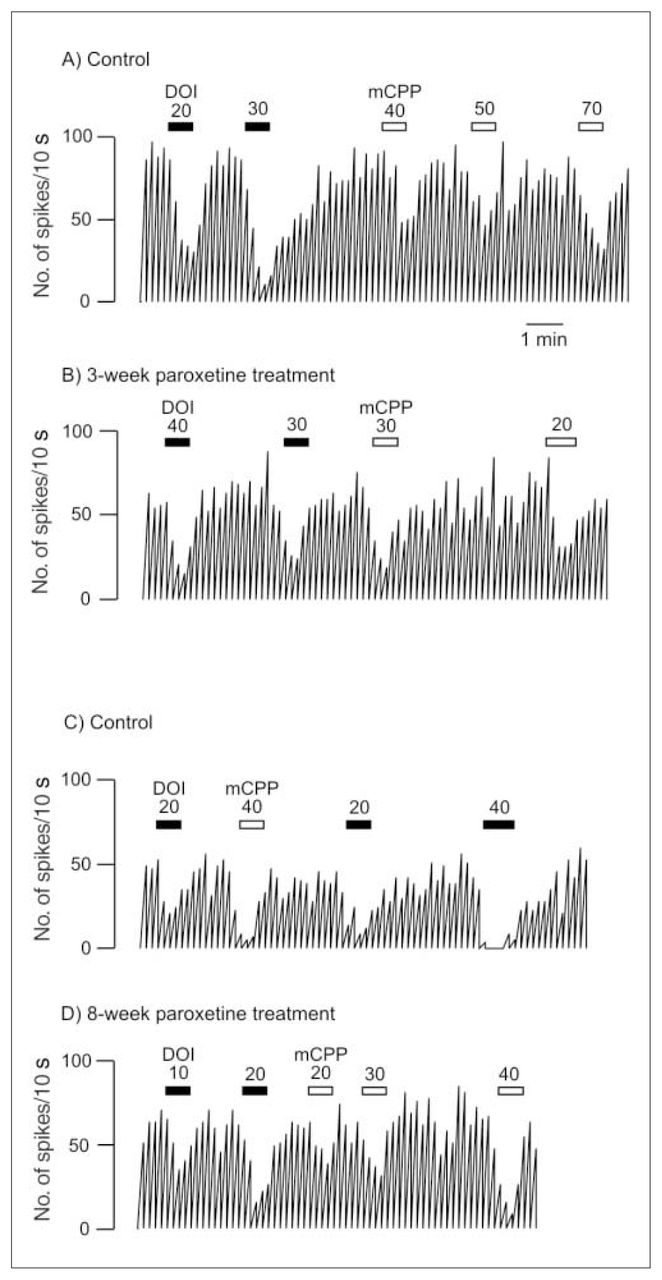
Integrated firing rate histogram of an orbitofrontal cortex neuron, showing its responsiveness to microiontophoretic application of DOI and mCPP in control animals and rats treated with paroxetine for 3 and 8 weeks. Horizontal bars indicate the duration of the applications (currents given in nanoamperes). The time base in A applies to all traces. Traces in the control and the treated rats were obtained from animals from the same shipment that were treated simultaneously after administration of vehicle or paroxetine. These traces were all obtained on the same day.
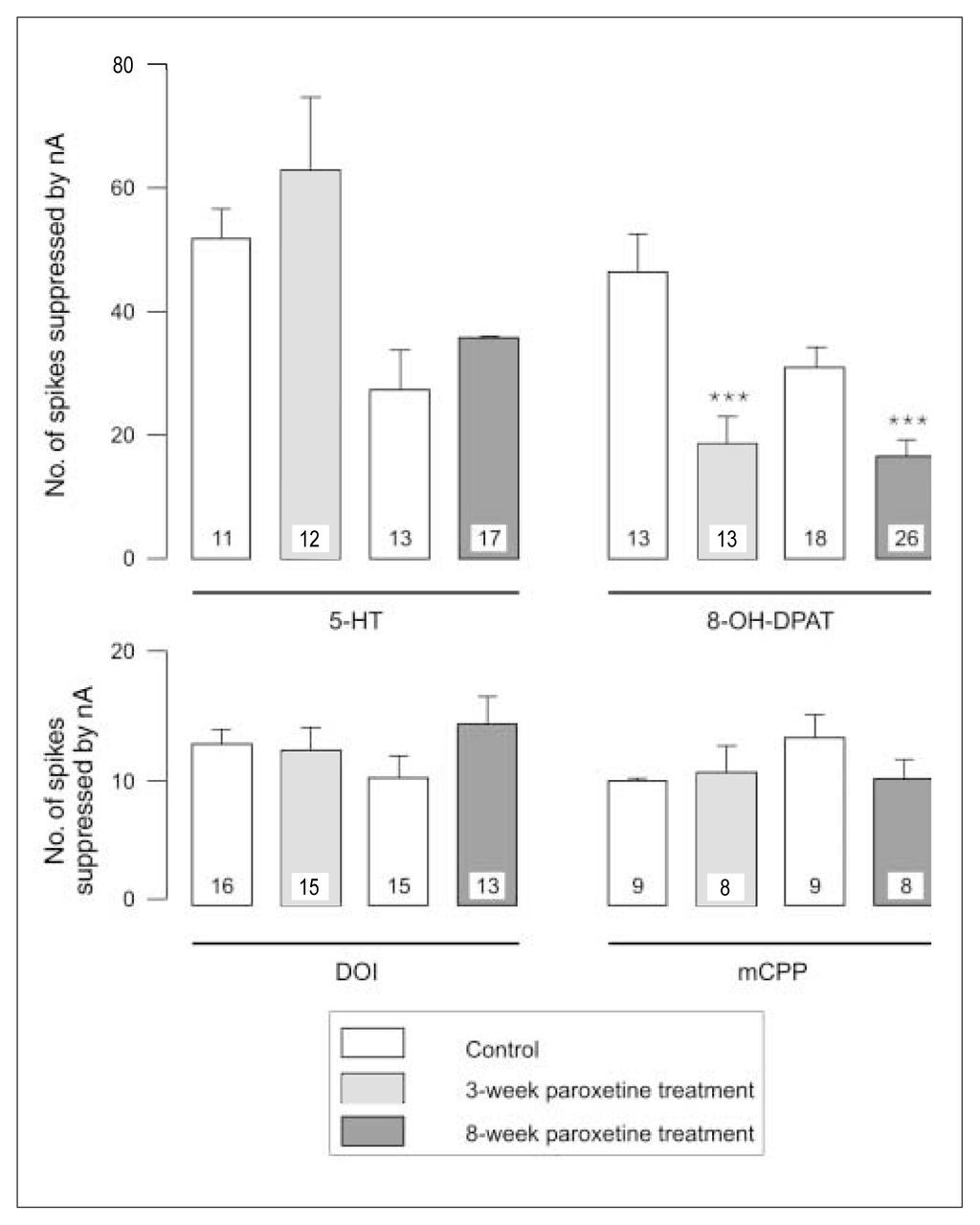
After 3 weeks of administration of paroxetine, there was no modification in the inhibitory effect of 5-HT, DOI or mCPP (Fig. 1, Fig. 2 and Fig. 3) in the OFC, whereas the inhibitory effect of 8-OH-DPAT was significantly attenuated (t24 = 4.05, p < 0.005; Figs. 1 and 3). This same attenuation of the inhibitory response of 8-OH-DPAT in the OFC was obtained after 8 weeks of paroxetine treatment (t42 = 5.18, p < 0.001; Fig. 1 and Fig. 3), whereas the responsiveness to 5-HT, DOI and mCPP remained unchanged (Fig. 1, Fig. 2 and Fig. 3).

Responsiveness to 5-HT, 8-OH-DPAT, DOI and mCPP in control animals and rats treated with paroxetine, expressed as the number of spikes suppressed by current (nA); the number in each column represents the number of neurons tested. *** p < 0.001.
It is noteworthy that the data for the 3-week treatment group were generated in part by assessment of neuronal responsiveness using the same micropipette in pairs of rats, a control rat and a treated rat taken from the same shipment, housed together and receiving the vehicle or the SSRI, respectively, for the same period of time. Use of the same micropipette in assessing neuronal responsiveness in such pairs of animals was designed to ascertain whether the difference, or lack of difference, between rats did not result merely from variations in the properties of the micropipettes. In 3 such pairs, 2 pipettes filled with 5-HT and 8-OH-DPAT or with mCPP and DOI were used, and the results from these experiments did not differ from the others.
Discussion
The present results show that sustained paroxetine administration, over 3 and 8 weeks, induced a decrease in the inhibitory effect of the 5-HT1A receptor agonist 8-OH-DPAT, which indicates a desensitization of postsynaptic 5-HT1A receptors in the OFC. Such desensitization had already been reported in at least 2 brain structures. In the amygdala, postsynaptic 5-HT1A receptor-mediated feedback, which regulates serotonergic neuronal firing rate in the dorsal raphe nucleus and 5-HT release, is diminished after 2 weeks of treatment with the SSRI citalopram.24 In addition, long-term treatment with paroxetine and fluoxetine reduces the neuroendocrine response to 8-OH-DPAT, which provides evidence of desensitization of hypothalamic postsynaptic 5-HT1A receptors.16–18 In vivo and in vitro electrophysiologic studies indicate, however, that in the hippocampus the sensitivity of postsynaptic 5-HT1A receptors is unchanged after administration of various SSRIs, including paroxetine, for 2–3 weeks.25,26 Interestingly, in constitutional 5-HT transporter knockout, a small but significant 5-HT1A receptor desensitization was observed in the hippocampus.27 Similarly, after a 3-week treatment with the selective monoamine oxidase type A inhibitor clorgyline, the responsiveness of pyramidal neurons to 5-HT itself, which is mediated by 5-HT1A receptors, is attenuated.28 Consequently this population of 5-HT1A receptors does have the capacity to desensitize following exposure to enhanced levels of synaptic 5-HT, but the desensitization does not occur with sustained administration of SSRIs.
The desensitization of 5-HT1A receptors in the OFC contrasts with results obtained by Bobula et al29 for the frontal cortex; in that study, the inhibitory effect of 8-OH-DPAT was enhanced in slices prepared from rats and treated with citalopram or imipramine for 14 days. It is noteworthy, however, that these experiments did not assess 5-HT1A receptor responsiveness under physiologic conditions, but rather under epileptic activity triggered by picrotoxin. Interestingly, the 5-HT1A receptor response to 8-OH-DPAT in the OFC appears atypical.22 Indeed, the inhibitory effect of 8-OH-DPAT was not blocked by the 5-HT1A receptor antagonists WAY 100635 or BMY7378. Also, the inhibitory effect of the 5-HT1A receptor agonist gepirone on OFC neurons was not antagonized by WAY 100635. Since 8-OH-DPAT but not gepirone is endowed with 5-HT7 receptor affinity, these observations have excluded a contribution of the latter receptor subtype to the action of 8-OH-DPAT and gepirone. These results would still be compatible with the existence of different 5-HT1A receptor subtypes or a different mechanism modulating the function of this receptor. A regional difference in the functional and regulatory properties of 5-HT1A sites has already been reported within the same brain structure, i.e., CA1 versus dentate gyrus in the hippocampus.30 This difference in properties is supported by the existence of 3 separate mRNA bands coding for 5-HT1A receptors in the rat brain.31
The desensitization of 5-HT1A receptors in the OFC could account for the lack of beneficial effect of the addition of the 5-HT1A receptor agonist buspirone to the regimen of patients with SSRI-resistant OCD.32,33 Indeed, it would be difficult for buspirone to increase the activation of 5-HT1A receptors in the OFC if the latter were desensitized. Consistent with this possibility is the report of Lesch et al34 showing that long-term exposure to fluoxetine attenuated the adrenocorticotropic hormone response to the 5-HT1A receptor agonist ipsapirone in patients with OCD; this finding suggests that long-term exposure to fluoxetine results in hyposensitivity of that 5-HT1A receptor function, in accordance with the desensitization of 5-HT1A receptors observed in the rat hypothalamus after long-term treatment with paroxetine and fluoxetine.16–18
In the presence of desensitized 5-HT1A receptors in the OFC, the inhibitory effect of 5-HT was unchanged in rats treated with paroxetine, which suggests that another 5-HT receptor may be mediating most of the effect of the endogenous neurotransmitter. Indeed, it is possible that 5-HT exerts its action via 5-HT2 receptors, since the present data show that these receptors are not desensitized after 3 and 8 weeks of treatment with paroxetine. These results contrast with earlier data showing attenuation of the excitatory effect on 5-HT2 receptors with application of DOI, based on recordings from slices of frontal cortex.29 Indeed, excitatory 5-HT2-mediated responses have generally been reported in in vitro experiments using bath application of agonists. It is thus possible that different populations of 5-HT2 receptors are activated in these experimental paradigms: 5-HT2 receptors located on both the cell body and the dendritic tree are activated when 5-HT agonists are used in a bath application, whereas 5-HT2 receptors located primarily on the cell body are activated when agonists are ejected by microiontophoresis. Because of a lack of variety of specific ligands, the exact receptor subtype mediating the effect of DOI and mCPP in the OFC remains controversial. Nevertheless, combinations of different strategies have led to the conclusion that 5-HT2 receptors in the OFC are pharmacologically distinct from those in other regions of the cerebral cortex. Studies from our laboratory have documented that the inhibitory effect of DOI and mCPP is attenuated by the 5-HT2 receptor antagonists ritanserin and risperidone in the medial prefrontal cortex but is not antagonized by ritanserin and the 5-HT1/2 antagonist metergoline in the OFC, indicating that 5-HT2 receptor responses may be mediated by an atypical receptor in the OFC of rats and guinea pigs.21,22 On the other hand, using 5-HT2C receptor null mutant mice, Rueter et al23 showed that DOI may be acting predominantly via a 5-HT2A receptor and that the effect of mCPP is mediated by both 5-HT2A and 5-HT2C receptors in the OFC.
If the antiobsessional effect of SSRIs depends on ongoing enhancement of serotonergic transmission, administration of a potent 5-HT antagonist might be expected to provoke symptom exacerbation in patients with a response to SSRIs. Indeed, the administration of the 5-HT1/2 antagonist metergoline over 4 days causes an exacerbation of OCD symptoms in patients with this condition.2,35 This result is in accordance with a report that the subacute administration of metergoline does attenuate the responsiveness of DOI in the caudate nucleus. 22 Consequently, attenuation of inhibitory 5-HT2 receptor neurotransmission at any point in the neuronal OCD circuitry, whether it be the 5-HT2 receptors in the caudate nucleus or those in the OFC, should reinstate the initial hyperactivity in the neuronal loop and trigger symptom exacerbation in patients with OCD whose condition has improved with SRI therapy.
Both 5-HT2A and 5-HT2C receptor antagonism have been postulated to play a role in the generation of obsessive–compulsive symptoms in patients with a psychotic disorder. Although low doses of the atypical antipsychotic risperidone have clearly shown a therapeutic effect in SSRI-resistant OCD,36,37 exacerbation of OCD with high doses of clozapine and risperidone may be due to the antagonism of 5-HT2 receptors in the OFC. Indeed, with low doses of risperidone, which block 5-HT2 receptor response in the medial prefrontal cortex, the responsiveness of 5-HT2 receptors in the OFC is unaffected. In contrast, high doses of the same drug attenuate the responsiveness of 5-HT2 receptors in the OFC.21
Consistent with the possibility that enhanced 5-HT2 receptor activation might underlie the beneficial action of SSRIs, case studies have reported that the use of hallucinogens with 5-HT2 receptor agonistic properties, such as LSD and psilocybin, results in relief of OCD symptoms.38–40 Moreover, it was demonstrated in an electrophysiology study that LSD enhances the inhibitory effect of 5-HT in the OFC, possibly through 5-HT2 receptors, but not in the hippocampus, where 5-HT exerts most of its action in these conditions via 5-HT1A receptors.41 Taken together, these clinical observations indicate that by enhancing the activation of 5-HT2 receptors with 5-HT agonists, a beneficial effect might be obtained in OCD, whereas by decreasing transmission at these sites with high doses of antagonists, OCD symptoms may be exacerbated in patients whose condition has previously been improved by an SSRI or produced de novo in patients with schizophrenia.
Acknowledgements
This work was supported by a Canadian Institutes for Health Research grant and by salary support from the University of Ottawa Institute of Mental Health Research to both authors, as well as a Research Chair in Psychopharmacology from the Canadian government to Dr. Blier.
Footnotes
Medical subject headings: serotonin; prefrontal cortex; receptor, serotonin, 5-HT1A; receptors, serotonin, 5-HT2; paroxetine; obsessive-compulsive disorder.
Competing interests: None declared.
Contributors: Drs. El Mansari and Blier conceived and designed the study, collected and interpreted the data, drafted and revised the article, and gave final approval for it to be published.
- Received January 26, 2005.
- Revision received March 28, 2005.
- Accepted April 11, 2005.
References

{kind=link}
{kind=link}
{kind=link}
Article tools






Related Articles
Cited By...
- No citing articles found.