

Abstract
Background: Increased oxidative stress is strongly implicated in bipolar disorder (BD), where protein oxidation, lipid peroxidation and oxidative damage to DNA have been consistently reported. High levels of dopamine (DA) in mania are also well-recognized in patients with BD, and DA produces reactive oxygen species and electron-deficient quinones that can oxidize proteins when it is metabolized.
Methods: Using immunohistochemistry and acceptor photobleaching Förster resonance energy transfer (FRET), we examined oxidation and nitration of areas immunoreactive for the DA transporter (DAT) and tyrosine hydroxylase (TH) in the postmortem prefrontal cortex from patients with BD, schizophrenia and major depression as well as nonpsychiatric controls.
Results: We found increased oxidation of DAT-immunoreactive regions in patients with BD (F3,48 = 6.76, p = 0.001; Dunnett post hoc test p = 0.001) and decreased nitration of TH-immunoreactive regions in both patients with BD (F3,45 = 3.10, p = 0.036; Dunnett post hoc test p = 0.011) and schizophrenia (p = 0.027). On the other hand, we found increased global levels of oxidation in patients with BD (F3,44 = 6.74, p = 0.001; Dunnett post hoc test p = 0.001) and schizophrenia (p = 0.020), although nitration levels did not differ between the groups (F3,46 = 1.75; p = 0.17).
Limitations: Limitations of this study include the use of postmortem brain sections, which may have been affected by factors such as postmortem interval and antemortem agonal states, although demographic factors and postmortem interval were accounted for in our statistical analysis.
Conclusion: These findings suggest alterations in levels of protein oxidation and nitration in DA-rich regions of the prefrontal cortex in patients with BD and schizophrenia, but more markedly in those with BD.
Introduction
Bipolar disorder (BD) is characterized by recurrent episodes of mania/hypomania and depression, affects 1.5% of the population, and is associated with high morbidity and mortality.1 The pathophysiology of BD is linked to a number of factors, including neurotransmitter imbalance, oxidative stress and genetic causes.2 Increased oxidative stress, which could result in oxidative and nitrosative damage to biomolecules,3 is a consistent finding in patients with BD. For example, mitochondrial dysfunction and decreased expression of genes of the electron transport chain, particularly that of complex I, are reported in patients with BD.4,5 Decreased efficiency of the electron transport chain could result in increased production of reactive oxygen species (ROS).3 Moreover, increased levels of carbonyl groups, 3-nitrotyrosine (3NT) and decreased levels of antioxidants, such as glutathione, are all reported in patients with BD.6–9
It is widely held that dysregulation of the dopamine (DA) system is important in BD, where high levels of DA are thought to underlie mania, a defining feature of the disorder.10 Increasing synaptic DA levels with amphetamine and Levodopa (L-dopa) produces mania-like behaviour,10,11 and antipsychotics, which act in part by blocking DA receptors, are among the most effective treatments for acute mania.12 High levels of DA can be cytotoxic in part through the generation of ROS and electron-deficient quinones during oxidation of DA.13 In fact, amphetamine increases markers of oxidative stress, such as lipid peroxidation and protein oxidation, in animals.11,14
Therefore, in this study, we examined oxidative and nitrosative damage in DA-rich regions of the prefrontal cortex (PFC) using acceptor photobleaching Förster resonance energy transfer (apFRET). We examined the PFC, as it was previously shown to have increased oxidative stress in patients with BD.5 In order to examine DA-rich areas, we labelled the dopamine transporter (DAT), which is a transmembrane protein responsible for the uptake of synaptic DA into the pre-synaptic terminal,15,16 and tyrosine hydroxylase (TH), which converts L-tyrosine to L-dopa and is the rate-limiting enzyme in DA synthesis. Previous studies used TH and DAT labelling with immunohistochemistry techniques to visualize dopaminergic axons in the PFC and reported extensive co-localization between TH- and DAT-immunoreactive (TH-IR/ DAT-IR) axons.15,16 Here, we report evidence of oxidative and nitrosative damage in DA-rich regions of the postmortem PFC of patients with BD and schizophrenia.
Methods
Postmortem brain samples
We obtained frozen postmortem PFC (Brodmann areas [BA] 9 and 46) sections (14 μm) from the Stanley Foundation Neuropathology Consortium (Table 1). The details of these samples have been published elsewhere.17 Briefly, the groups consisted of samples from patients with BD, major depressive disorder (MDD) and schizophrenia and from nonpsychiatric controls. There were no differences in sex (F3,55 = 0.11, p > 0.99), age (F3,55 = 0.53, p = 0.67), postmortem interval (PMI; F3,55 = 1.70, p = 0.18) and pH (F3,55 = 0.63, p = 0.60) among the groups (n = 15 per group). Diagnoses were established using DSM-IV criteria. We used sections with good structural integrity. Samples were randomly coded numerically, and the experimenters were kept blind to the codes until all the experiments were completed.
Demographic and clinical characteristics of sample by group
Immunohistochemistry
We Nissl stained and labelled sections for NeuN to examine tissue quality before immunohistochemistry experiments (Fig. 1A and 1B; for supplementary methods see the Appendix, available at jpn.ca). Four adjacent sections per donor were processed for immunohistochemistry, where 2 sections were labelled for DAT and free thiols and the other 2 were labelled for TH and 3NT. We performed the FRET analysis and intensity analysis simultaneously on the same sections. 7-Diethylamino-3-(4′maleimidylphenyl)-4-methylcoumarin (CPM; Invitrogen; D346) is a specific label for free thiols.18 We completed CPM labelling using a previously published method by Mastroberardino and colleagues18 with slight modifications. We used CPM to directly label free thiols without prior modification of the tissue to minimize alterations to the sections and introduction of modifications from the experimental procedure. The indirect method by Mastroberardino and colleagues, in which free thiols are blocked and disulfide linkages are reduced to be labelled with CPM, was used to increase sensitivity.18 For our samples, the direct labelling method was sufficient to detect between-subject differences. In the methods described here, lower CPM labelling indicates greater oxidation of thiols. Sections fixed with 4% para-formaldehyde (10 min) were incubated in 0.5 mM of CPM in Tris-HCl (0.1 M, pH 6.8) for 1 hour. Sections were washed extensively, and we carried out immunohistochemistry using previously published techniques.19 Sections were blocked in 4% goat serum for 1 hour and incubated with the DAT antibody (Millipore; MAB369, 1:250, 60 h at 4°C) in 0.05% Triton-X. The specificity of this antibody has been demonstrated in previous studies.20,21 Sections were then washed, incubated with Alexa Fluor®488 antirat (Molecuar Probes, A11006) in 0.05% Triton-X for 3 hours and mounted in Fluoromount™ (Cedarlane). To create a negative control for CPM labelling, paraformaldehyde-fixed sections were alkylated with 100 mM N-ethylmaleimide (Bioshop; ETM222) and 100 mM iodoacetamide (Sigma-Aldrich; I6125) in Tris-0.1 M, pH 6.8 (15 min) before CPM labelling. Oxidation of thiol groups with N-ethylmaleimide and iodoacetamide diminished CPM labelling, demonstrating specificity of CPM for free thiols. Hence, decreased CPM labelling indicates greater thiol oxidation. These sections were used as controls to account for nonspecific labelling of CPM.18 For 3NT and TH double labelling, acetone-fixed sections were blocked with 10% goat serum in 0.3% phosphate-buffered saline Triton X-100 (PBS-T) for 30 minutes. Sections were then incubated with primary antibodies in PBS-T (48 h, 4°C): TH (Millipore; AB152, 1:250) and 3NT (Abcam; ab61392, 1:200). Sections were washed, incubated with secondary antibodies (2 h, room temperature) in PBS-T (1:300): Alexa Fluor®488 antirabbit immunoglobin G (Molecular probes®; A11008) and Alexa Fluor®350 antimouse immunoglobin G (Molecular probes®; A11045) and mounted in FluoromountTM (Cedarlane). Specificity of the anti-TH antibody has been demonstrated elsewhere.22 The anti-3NT antibody was previously used by our laboratory.7 To test the specificity of the anti-3NT antibody, sections were treated with peroxynitrite (Millipore, 20–107, 1:100), which is used as a positive control for 3NT labelling, or degraded peroxynitrite (Millipore, 20–247, 1:100) using a previously published method.23 An increase in 3NT labelling was observed only with peroxynitrite, demonstrating the specificity of the anti-3NT antibody (Appendix, Fig. S1). To minimize the risk of antibody cross-reactivity, different combinations of primary and secondary antibodies were tested, where only minor and nonspecific binding was found, indicating minimal cross-reactivity (Fig. 1C and 1D).

(A) Example of a Nissl-stained section taken with Nikon Eclipse 80i Microscope with good section quality. Scale = 44 μm. (B) Example of a section labelled for NeuN (green). Image was taken with a Nikon Eclipse 80i Microscope. Scale = 58 μm. (C) Example of a section labelled for dopamine transporter (DAT; green). Image was taken with a confocal laser scanning microscope (CLSM). The inset is an example of a section labelled with only the secondary antibody, demonstrating minimal labelling, and therefore specificity of the secondary antibody. (D) Example of a section labelled with the tyrosine hydroxylase (TH) antibody (green) taken with a CLSM. The inset is an example of a section labelled with the primary antibody for 3-nitrotyrosine (3NT), and secondary antibody for TH. Minimal labelling was observed, indicating specificity of the secondary antibody. (E) Example of a Förster resonance energy transfer (FRET) experiment for DAT and 7-diethylamino-3-(4′maleimidylphenyl)-4-methylcoumarin (CPM) FRET. The red circle indicates the area that has been photobleached. Increase in the intensity of the donor fluorophore (blue; CPM) can be observed after bleaching the acceptor fluorophore (Alexa488; green), indicating the presence of FRET. (F) Example of a FRET experiment for TH and 3NT FRET. The red circle indicates the photobleached area. Increase in the intensity of the donor fluorophore (Alexa350; blue) can be observed after photobleaching the acceptor fluorophore (Alexa488; green), indicating presence of FRET.
Intensity analysis
Intensity analysis was performed using a previously published method from our group.24 We used a 60× water immersion objective and a confocal laser scanning microscope (CLSM; Olympus Fluoview FV1000, Olympus America Inc.) to capture images from 5 fields of each section, which were chosen by dividing the section into 5 equally sized rectangles, and imaging the middle region of each of the 5 rectangles. For CPM- and DAT-labelled sections, CPM was excited with the 405 nm laser (15% intensity) and Alexa488 was excited with the 488 nm laser (18% intensity). For 3NT- and TH-labelled sections, Alexa488 was excited with a 488 nm laser (15% laser intensity) and Alexa350 was excited with a 405 nm laser (5% intensity). Detection wavelength for Alexa488 was 500–600 nm and for Alexa350/CPM was 425–475 nm. Confocal aperture diameter was 110 μm, and line-by-line sequential detection mode was used. Scanning parameters were consistent for all experiments. We performed image analysis using the FV10-ASW software version 2.1 (Olympus). For analyzing DAT, TH and 3NT intensity, we first subtracted background from the image by selecting 5 regions per field without specific immunoreactivity as background and measuring the average intensity of the regions. This value was subtracted from the overall intensity of the field. We then calculated the average intensity of the 5 fields to represent the intensity for the section. Therefore, DAT/TH/3NT intensity = (average intensity of 5 fields from the section) – (average background intensity of 5 fields from the section). We determined CPM intensity using the sections treated with iodoacetamide and N-ethylmaleimide (negative control) to correct for nonspecific labelling,18 which had significantly decreased CPM labelling, demonstrating its specificity for free thiols. Therefore, CPM intensity = (average intensity of 5 fields from the section) – (average intensity of 5 fields from negative control).
Förster Resonance Energy Transfer
Acceptor photobleaching FRET measures nonradiative energy transfer from a donor fluorophore to an acceptor fluorophore by observing the fluorescence of the donor in the presence and absence (photobleaching) of the acceptor.25 Acceptor photobleaching FRET has been used successfully in previous studies with human samples.25–27 The energy transfer between a donor and an acceptor fluorophore occurs only when the donor and the acceptor are within about 30–50 nm of each other using indirect immunohistochemistry, which allows us to use this technique to detect the proximity of the molecules.25 For DAT and CPM FRET, Alexa488 (DAT) was the acceptor fluorophore and CPM was the donor fluorophore. For 3NT and TH FRET, Alexa488 (TH) was the acceptor fluorophore and Alexa350 (3NT) was the donor fluorophore. We performed apFRET with a CLSM (Olympus) using previously published techniques with small modifications.18,25,26 We used a 60× water immersion objective. Image acquisition parameters were identical to those used for intensity analysis. We measured the intensity of the donor fluorophore before and after the photobleaching of the acceptor fluorophore. Photobleaching of the acceptor fluorophore was at 100% laser intensity (10 μs/pixel) using the 488 nm laser in a single bleaching step (1.5 s for TH and 3NT, 60 ms for DAT and CPM). Scanning parameters were consistent during all experiments. Change in fluorescence was represented as increase in fluorescence (ΔIF), where ΔIF = intensity of donor after photobleaching – intensity of donor before photobleaching. To account for non-specific binding of CPM, we performed negative controls where sections were treated with iodoacetamide and N-ethylmaleimide before CPM staining and immunohistochemistry as detailed previously. Then we subtracted the ΔIF of the negative controls from the ΔIF determined for the samples to calculate the final ΔIF.18 The ΔIF between CPM and the secondary antibody (Alexa488) without the primary antibody against DAT was undetectable. To test the effect of antibody cross-reactivity on FRET measurements for 3NT and TH FRET, we labelled sections with only the primary antibody for TH and both secondary antibodies, then we measured FRET. A very low level of FRET was detected (average ΔIF = 12.5), indicating negligible cross-reactivity. Five regions of interest (ROI) per section were chosen as described in the intensity analysis section and examined. Therefore, ΔIF = (average ΔIF of 5 ROIs in section) – (average ΔIF of 5 ROIs in negative control). We used the ΔIF to measure energy transfer, which was demonstrated to be more appropriate with tissue sections than FRET efficiency owing to its lower sensitivity to non-specific binding of fluorophores.25 For DAT and CPM FRET, lower ΔIF indicates lower levels of free thiols in DAT-IR regions, and thus likely increased oxidative stress status in DAT-IR regions. For TH and 3NT FRET, higher ΔIF indicates greater 3NT levels in TH-IR regions. All calculations were performed using FV10-ASW software version 2.1.
Mouse brain study for the effect of PMI on oxidation of neurons
We used adult male C57/BL6 mice (8–12 wk) to examine the effect of PMI on thiol oxidation and 3NT formation in neurons. The University Animal Care Committee of the University of Toronto approved our study protocol (protocol number 20009477), and care of animals was performed according to animal research guidelines of the University of Toronto. Animals were euthanized by asphyxiation and kept at 4°C until an appropriate PMI was reached (6, 24, 48, 72 h) to mimic the storage of cadavers.28 The brain was then extracted, sagittally bisected, flash frozen in isopentane and dry ice, and stored at −80°C.17 Each bisected brain was processed and analyzed independently. Sagittal sections of 14 μm were made using a cryostat, and using standard immunohistochemistry techniques, acetone- and methanol-fixed sections were labelled with NeuN (Millipore; MAB377B) and CPM or 3NT (abcam; ab61392) and NeuN. The secondary antibodies were streptavidin-Alexa Fluor®568 (Molecular probes®; S11226) and Alexa Fluor®350 anti-mouse immunoglobulin G (Molecular probes®; A11045). Controls for CPM and 3NT labelling were carried out as described above. Sections were then used to obtain images of the frontal cortex with a CLSM (Olympus) using a 60× water immersion objective. The CPM and Alexa350 were excited with the 405 nm laser (32% intensity) and Alexa568 was excited with the 543 nm laser (38% intensity). Detection wavelength for Alexa568 was 555–655 nm and for CPM and Alexa350 was 425–475 nm. The images were analyzed using colocalization analysis (Pearson correlation coefficient) between NeuN and CPM (free thiols in neurons) or between NeuN and 3NT (protein nitration in neurons) with FV10-ASW software version 2.1.
Statistical analysis
We performed statistical analyses using SPSS software version 20. We determined normal distribution of data using the Kolmogorov–Smirnov test. As the data were normally distributed, we used parametric tests for further analysis. We assessed the effect of covariates (age, sex, PMI and pH) using the Pearson correlation test. As age was the only variable that significantly correlated with one of our dependent variables (DAT intensity; Pearson r = 0.34, p = 0.031), we used an analysis of covariance (ANCOVA) model with age as a covariate and performed a Dunnett post hoc test to examine the effect of diagnosis. Data are presented as mean ± standard error of the mean (SEM).
Results
Sample characteristics
Our sample consisted of patients with BD (n = 15), schizophrenia (n = 15) and MDD (n = 15) and of nonpsychiatric controls (n = 15). Demographic and clinical characteristics of the sample are summarized in Table 1. Age, PMI and brain pH did not significantly correlate with CPM intensity, 3NT immunoreactivity, TH immunoreactivity, ΔIF between DAT and CPM and ΔIF between 3NT and TH (Fig. 2). Dopamine transporter immunoreactivity did not correlate with PMI or pH but positively correlated with age (Pearson r = 0.34, p = 0.031). Furthermore, psychiatric patients with a history of substance abuse had lower DAT intensity (F2,46 = 6.74, p = 0.003) than nonpsychiatric controls (p = 0.001). We Nissl stained the sections to examine their quality and found that sections had good histological quality without large holes or tears (Fig. 1A). Also, the average glia:neuron ratio (1:37) was comparable to that published in previous studies examining the same area.29,30

(A) Correlation between ΔIF for dopamine transporter (DAT) and 7-Diethylamino-3-(4′maleimidylphenyl)-4-methylcoumarin (CPM) and brain pH, age and postmortem interval (PMI). (B) Correlation between ΔIF for tyrosine hydroxylase (TH) and 3-nitrotyrosine (3NT) and brain pH, age and PMI. Results were assessed using the Pearson correlation coefficient. The correlations were not significant.
Dopamine transporter and TH immunoreactivity
We examined DAT and TH immunoreactivity in the sections by measuring fluorescence intensity. The groups did not differ for DAT immunoreactivity (F3,44 = 1.48, p = 0.23; Fig. 1C) or TH immunoreactivity (F3,46 = 0.86, p = 0.47; Fig. 1D).
Intensity of CPM and 3NT immunoreactivity
We quantified the intensity of CPM to assess the amount of free thiols in the sections. There was a significant difference among the groups (F3,44 = 6.74, p = 0.001; Fig. 3A), where patients with BD (p = 0.001) and schizophrenia (p = 0.020) had lower CPM intensity in the PFC than nonpsychiatric controls. Age, sex, PMI and pH did not correlate with CPM or 3NT intensity. To explore the potential effect of psychotropic drugs, we compared psychiatric patients who were taking lithium, antipsychotics or antidepressants at the time of death with patients who were taking drugs and with nonpsychiatric controls. Patients who were taking antipsychotics at the time of death were found to have lower CPM intensity (F2,44 = 7.69, p = 0.001) than controls (p = 0.001). On the other hand, patients who were not taking antidepressants had lower CPM intensity (F2,44 = 8.31, p = 0.001) than controls (p = 0.001). There were no other effects of drug treatment on either CPM intensity or 3NT immunoreactivity. We also assessed 3NT immunoreactivity by measuring fluorescence intensity. In contrast to CPM intensity, 3NT immunoreactivity did not differ among the groups (F3,45 = 1.80, p = 0.16; Fig. 3B). Because protein stability may be affected by longer PMIs, we examined the effect of longer PMIs on the frontal cortex using mouse brain. We examined neurons as they have lower antioxidant capacity than glial cells.3 We found that CPM (F3,12 = 1.30, p = 0.32) and 3NT labelling (F3,12 = 0.82, p = 0.82) in neurons did not change with increasing PMI up to 72 hours.
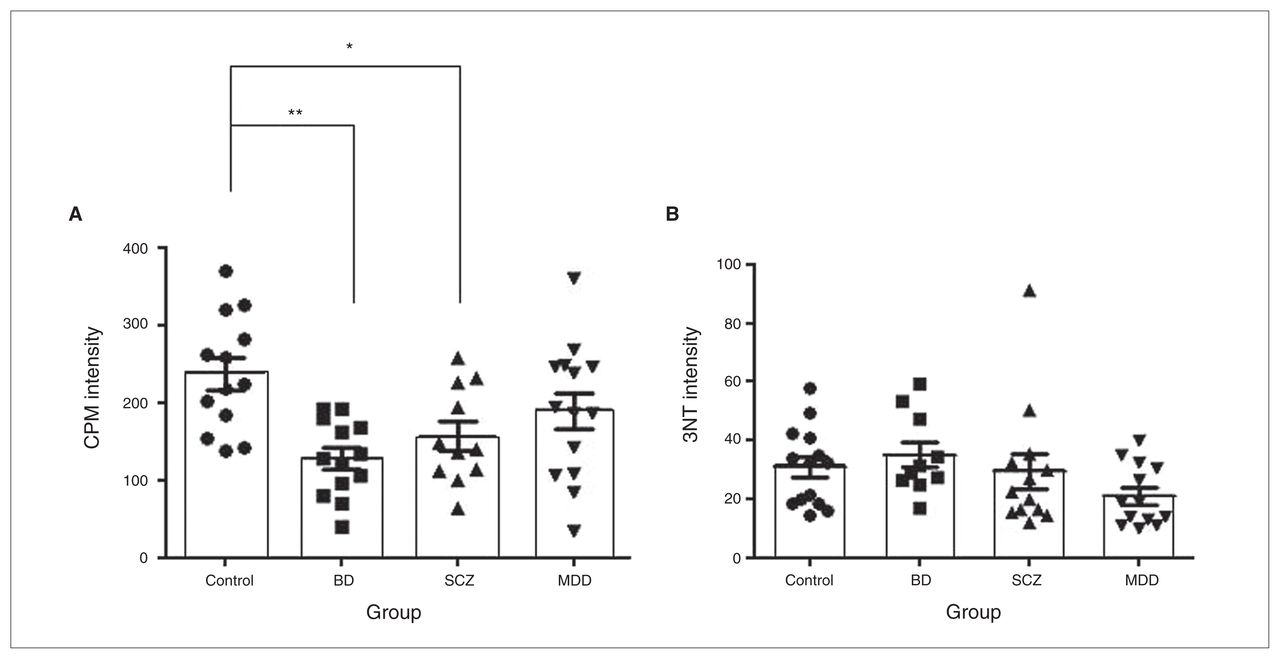
(A) Intensity of 7-Diethylamino-3-(4′maleimidylphenyl)-4-methylcoumarin (CPM) labelling after correction for nonspecific CPM binding with negative control (sections treated with iodoacetamide and N-ethylmaleimide). Results are expressed as mean ± standard error of the mean (SEM). *p < 0.05; **p < 0.01. n = 50 (control, n = 13; bipolar disorder [BD], n = 13; schizophrenia [SCZ] = 11, major depressive disorder [MDD] = 13). (B) 3NT immunoreactivity after correction for nonspecific signals as measured by fluorescence intensity. Results are expressed as mean ± SEM, n = 50 (control = 14, BD = 10, SCZ = 13, MDD = 13). All analyses were performed on nonprocessed, original images in original imaging format (OIF).
Acceptor photobleaching FRET analysis
Initially, our aim was to examine oxidation and nitration in both TH-IR and DAT-IR regions using FRET analysis. However, FRET between CPM and Alexa488 labelling TH was not detectable in our sections. Moreover, FRET between Alexa350 labelling 3NT and Alexa488 labelling DAT was not detectable. Therefore, we examined free thiols in DAT-IR regions (DAT and CPM FRET) and nitration of TH-IR regions (TH and 3NT FRET). There was a significant difference among the groups for DAT and CPM FRET (F3,48 = 6.76, p = 0.001), where patients with BD had lower energy transfer between CPM and DAT (Alexa488) than controls (p = 0.001, Fig. 4A). Also, the groups differed significantly for TH and 3NT FRET (F3,45 = 3.10, p = 0.036; Fig. 4B), where patients with BD (p = 0.011) and schizophrenia (p = 0.027) had lower levels of nitration than controls. Similarly, to explore the potential effect of psycho-tropic drugs, we compared patients who were prescribed antipsychotics, antidepressants or lithium with those who were not prescribed drugs and also with nonpsychiatric controls. The only significant effect was that patients who were not prescribed antidepressants had lower TH and 3NT FRET (F2,42 = 3.26; p = 0.048) than healthy controls (p = 0.027), although this effect was not observed when comparing those who were prescribed antidepressants with controls.

(A) Change in fluorescence (ΔIF) between Alexa488 labelling dopamine transporter (DAT) and CPM labelling free thiols. Higher values indicate greater amounts of free thiols in DAT-IR regions after correction for background ΔIF (FRET in oxidized sections). Results are expressed as mean ± SEM *p < 0.05 **p < 0.01. n = 54 (Control = 14, bipolar disorder [BD] = 14; schizophrenia [SCZ] n = 12; major depressive disorder [MDD] n = 14). (B) ΔIF between Alexa488 labelling tyrosine hydroxylase (TH), and Alexa350 labelling 3-nitrotyrosine (3NT). Greater ΔIF indicates greater nitration. Results are expressed as mean ± SEM. *p < 0.05; **p < 0.01. n = 50 (Control n = 11, BD n = 11, SCZ n = 14, MDD n = 14). All analyses were performed on nonprocessed, original images in Original Imaging Format (OIF).
Discussion
In this study, we examined the association between oxidative stress and DA in patients with BD by measuring oxidative and nitrosative damage in DA-rich areas of the PFC. Using FRET, we found increased oxidation of DAT-IR areas in patients with BD, whereas nitration of TH-IR areas was decreased in patients with BD and schizophrenia. When we examined global levels of oxidative and nitrosative damage, however, we did not find between-group differences in levels of nitration, but found thiol oxidation to be increased in both patients with BD and schizophrenia. These results suggest alterations in oxidation and nitration of DA-rich areas in the PFC that are particularly marked in patients with BD, but also in those with schizophrenia.
When oxidation and nitration of DAT and TH immuno-reactivity were examined using FRET, we found increased oxidation in DAT-IR areas in patients with BD while nitration in TH-IR areas was decreased in both patients with BD and schizophrenia. These results suggest oxidative and nitrosative modifications to DA-rich areas in both patients with BD and schizophrenia, which could contribute to the dysregulation of the DA system hypothesized to occur in patients with these disorders. More specifically, hyperactivity of the DA system in mania is well-recognized, where antipsychotics, which act in part by blocking DA signalling, are among the most effective treatments for acute mania,12 and dopaminergic agonists have been found to produce mania-like behaviour in humans and in animals.10,31 The DA hypothesis for schizophrenia is more complex, however, as the direction of DA dysregulation may differ depending on symptoms (positive v. negative) and brain region (cortical v. subcortical).32 However, owing to the limitations of using postmortem brain sections, future studies examining the consequences of these alterations to DA signalling and behaviour are required. Furthermore, it is interesting to note that patients with BD and schizophrenia had similar alterations in our study, which is consistent with the results of previous studies showing extensive overlap between these 2 disorders, including cognitive deficits,33 genetic alterations,34 changes in brain morphology35 and oxidative stress.36 Our results also showed that patients who are not prescribed antidepressants have lower TH and 3NT FRET than controls. This effect is difficult to interpret. While antidepressants have been shown to have neuroprotective effects through mechanisms other than oxidative stress, including regulation of neurotrophin levels,37 it is not known whether the reduction in oxidative stress in patients treated with anti-depressants reflect their neuroprotective effects. A larger issue relates to the difficulty in disentangling the effects of medication and diagnosis, since antidepressants in most cases were prescribed to patients with depression but not to those with BD or schizophrenia.
Acceptor photobleaching FRET has a resolution of about 30–50 nm, and the effectiveness of FRET increases as the distance between the fluorophores decreases,25 suggesting that oxidation and nitration in DAT-IR and TH-IR areas as measured by FRET may in large part include direct oxidation of DAT and nitration of TH. Previous studies have demonstrated that DAT is a target of thiol oxidation but not nitration,38 which may have contributed to our difficulty measuring FRET between the fluorophores labelling DAT and 3NT (data not shown). As DAT oxidation decreases its ability to uptake DA,31,38 increased DAT oxidation in patients with BD may contribute to increased levels of synaptic DA. Whether TH is primarily a target of nitration or cysteine oxidation is controversial, with some studies showing evidence for both39 and others reporting TH nitration and oxidation only when the protein is unfolded.40,41 In the present study, we could not detect FRET between CPM labelling free thiols and TH. Since it is physiologically unlikely that all the cysteine residues of TH were in the oxidized state, we believe that CPM may not have been able to access the thiol groups of the TH protein as they are highly shielded from the surrounding environment.42 Decrease in 3NT levels in TH-IR regions was an unexpected finding, as increased oxidative stress is strongly implicated in BD.7 This finding may be specific to the TH protein or to L-dopa–rich areas, since DA, L-dopa and 3,4-dihydroxyphenylacetic acid are favourable substrates for reactive nitrogen species, and may protect proteins in their proximity that are less favourable substrates for nitration.39 Further study is required to elucidate this hypothesis. Also, because TH nitration has been shown to decrease its ability to produce L-dopa,39,41 decreased nitration of TH in patients with BD and schizophrenia suggests that nitration of TH may be present in normal conditions to limit the activity of this enzyme. Consequently, decreased nitration of TH in patients with BD and schizophrenia could act to “disinhibit” TH and result in higher levels of presynaptic DA. Also, while TH-IR and DAT-IR fibres colocalize in the human PFC,15 TH is also present in noradrenergic neurons, suggesting that decreased nitration in TH-IR regions may also occur in noradrenergic cells. Interestingly, dysregulation of noradrenergic signalling has also been shown in patients with BD,43 suggesting a potential role for decreased TH nitration in the noradrenergic system. Furthermore, while the focus of this study was the DA system, such changes could also occur in other neuronal populations. Future studies examining these alterations in different neuronal cells may increase our understanding of the pathophysiology of BD and schizophrenia.
Alterations in the PFC are consistently implicated in BD and other psychiatric disorders, where studies have reported decreased metabolism, morphological changes44 and increased oxidative damage.5 Moreover, the PFC has a high relative density of dopaminergic axons compared with other cortical regions,15 and DA is involved in the regulation of working memory by the PFC,45 which is impaired in patients with BD and schizophrenia.46,47 These studies suggest that dysregulation of the DA system in the PFC may be involved in the pathophysiology of these disorders. In agreement with previous studies, we found that TH and DAT labelling reveals fibres that may be dopaminergic axons. We also saw punctate structures that were occasionally connected by fibres, which may be axonal varicosities and terminals15,16 (Fig. 1C and Fig. 1D). Dopamine transporter and TH immunoreactivity examined by measuring fluorescence intensity did not differ among the groups, which is in agreement with the results of previous studies examining the same samples as those used in the present study (data can be found in the Stanley Neuropathology Consortium Integrative Database: http://sncid.stanleyresearch.org). We also found that DAT intensity was lower in patients with a history of substance abuse than in controls. Although the substances used by the patients are unknown, this finding suggests that drug abuse may influence the dopaminergic system in the PFC. It should, however, be noted that fluorescence intensity is not a truly quantitative measure of dopaminergic innervation. As our purpose was to visualize DA-rich areas of the PFC and our sections (frozen, 14 μm thick) were not amenable to stereological analysis,48 we did not pursue this further using more quantitative methods.16,48
Global levels of free thiols and 3NT were examined by measuring fluorescence intensity. We used CPM to label free thiols using a method validated by Mastroberardino and colleagues.18 We chose to use thiol oxidation as a marker of protein oxidation as it occurs more readily than formation of protein carbonyls and hence may be a more sensitive measure.18 3-Nitrotyrosine is a marker of protein nitration that has been used in other studies examining oxidative stress.5–7 We found increased thiol oxidation in patients with BD and schizophrenia, but we did not find between-group differences in protein nitration. Previous studies have consistently reported increased markers of oxidative stress for both patients with schizophrenia and BD, where increased levels of protein carbonyl groups and lipid peroxidation have been found.6,9 Oxidation of thiol groups can alter protein structure and function49 and may be a contributing factor in the pathophysiology of these disorders. Increased thiol oxidation in patients with BD and schizophrenia may occur owing to a number of different reasons, including lower levels of glutathione8,9 and mitochondrial dysfunction,4 particularly that of complex I, which could increase the production of ROS3 that can oxidize proteins. Decreased levels of expression of subunits of the mitochondrial electron transport chain have been reported for both patients with BD and schizophrenia.4 Since cell loss and morphological alterations are found in the PFC in patients with BD and schizophrenia,50–54 cellular dysfunction produced by protein oxidation may contribute to these alterations. This may lead to a decline in the proper functioning of the PFC. As mentioned previously, decreased working memory is consistently reported in patients with BD,55,56 suggesting that cellular dysfunction produced by oxidative damage could be contributing to this as well. Furthermore, we found an effect of prescribed medications, where patients who were prescribed antipsychotics had greater oxidative damage than controls. Since antipsychotics are D2 receptor antagonists, the effect of these drugs on oxidative damage in the DA system is of potential interest.31 Also, patients who were not prescribed antidepressants had greater global oxidative damage than those who were prescribed antidepressants, which is consistent with a potential protective effect of these drugs, as described for TH and 3NT FRET results. 3-nitrotyrosine, which is formed when reactive nitrogen species, such as peroxynitrite, react with tyrosine residues,57 was used as a marker of protein nitration. We chose to label 3-nitrotyrosine to examine protein nitration as it has been successfully used by our group and others to examine oxidative stress in postmortem brain samples.5–7 Although we did not find global 3NT levels to differ among the groups, a previous study showed increased 3NT levels in the PFC of patients with BD and schizophrenia.5 The difference between these findings may be explained by the use of different techniques, as the earlier study5 measured 3NT levels using the ELISA technique from whole tissue homogenates whereas the present study used immunohistochemistry to label cells in tissue sections. Moreover, the earlier study examined BA 10 whereas the present study examined BA 9 and BA 46. Brodmann areas 9 and 10 have different cytoarchitecture, suggesting that they may differ in the type and abundance of cells.58 Since different cells have different vulnerabilities to oxidative stress, this may have contributed to the difference between these 2 areas.3 Nonetheless, the differences in overall 3NT levels among diagnostic groups were in the same direction as those reported earlier by our group.5 Also, while global 3NT levels did not differ between groups, nitration of TH-IR regions measured by FRET was decreased in patients with BD and schizophrenia. This suggests that nitration may be a more specific phenomenon where increase or decrease in nitration of certain proteins does not always correspond with global levels of oxidative stress.57
Limitations
Our findings must be interpreted in light of their limitations. First, we had a small sample size and used multiple comparisons for statistical analysis. Second, we measured fluorescence intensity to compare DAT, TH and 3NT immunoreactivity among the groups. Fluorescence intensity obtained from tissue sections labelled with antibodies is a relative measure and is limited by factors such as photo-bleaching and tissue quality. To minimize the effect of these factors on our results, we kept the scanning parameters consistent, subtracted background intensity and used only sections with good histological quality. Also, because we used human postmortem brain samples, pre- and postmortem factors, such as antemortem agonal states, may have affected our results.59,60 To account for this, we examined the effect of demographic factors and postmortem alterations in data analysis by correlating our findings with PMI, pH and age. Furthermore, in agreement with the results of previous studies that examined PMIs of up to 48 hours,60,61 thiol oxidation and nitration in the mouse frontal cortex did not change up to a PMI of 72 hours. Although we found an effect of the history of substance abuse on 1 measure, DAT intensity, we do not have detailed information on the history of substance abuse or dependence to further explore how these factors could have influenced our results. In addition, while the effects of drug treatment on CPM intensity (anti-psychotics and antidepressants) and TH and 3NT FRET (antidepressants) are of potential interest, there are several important limitations. These include small sample size, the differences in prescribing patterns across diagnosis and absence of information on whether the patients were taking the prescribed medications. Other approaches not solely relying on postmortem brain samples may be more helpful to determine the effects of drug treatment on these measures. In addition, we examined only the PFC. Examining different areas of the brain may help to further elucidate the association between DA and oxidative stress in patients with BD and schizophrenia. Finally, use of other techniques, such as mass spectrometry to examine oxidative and nitrosative modifications to DAT and TH in postmortem brain and functional assays to examine the cause and consequences of such modifications may help elucidate the association between oxidative stress and DA in patients with BD and schizophrenia.
Conclusion
Our results demonstrate increased protein oxidation in the PFC of patients with BD and schizophrenia. We also found increased thiol oxidation in DAT-IR regions of patients with BD and dysregulation of protein nitration in TH-IR regions of patients with BD and schizophrenia, suggesting oxidative and nitrosative modifications to DA-rich areas in both of these disorders. Therefore, our findings suggest that the interaction between oxidative stress and DA may be important for the pathophysiology of BD and schizophrenia, and that future studies examining the role of oxidative modifications on dopaminergic proteins may contribute to a better understanding of how dysregulation of the DA system occurs in patients with these disorders.
Acknowledgements
We thank Ginnie Ng for her contributions in cell counting of the Nissl stained sections and Dr. Dennis Grant for donating the mice for the postmortem interval experiment. We also declare CIHR, Brain and Behaviour Research Foundation (NARSAD), and OGS as sources of funding, and Stanley Foundation Neuropathology Consortium as source of human postmortem brain sections.
Footnotes
Competing interests: None declared.
Contributors: H.K. Kim, A.C. Andreazza and L.T. Young designed the study. H.K. Kim, D.P.Y. Yeung and C. Isaacs-Trépanier acquired the data, which H.K. Kim, A.C. Andreazza, D.P.Y. Yeung and L.T. Young analyzed. H.K. Kim, A.C. Andreazza and L.T. Young wrote the article, which all authors reviewed and approved for publication.
- Received July 29, 2013.
- Revision received October 29, 2013.
- Revision received November 29, 2013.
- Accepted December 2, 2013.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Article tools





