

Abstract
Optimal early development in most species is dependent upon a stable relationship between the mother and her infant. The research described here focuses on the reciprocal nature of this dyad in rodents and humans, with respect to the regulation of responsiveness to stress in both mother and offspring. Dietary influences are critical not only to regulate infant growth but also to modulate the response of the neuroendocrine system to stress and, possibly, to influence some aspects of brain development. In particular, we discuss the role of leptin, a protein produced in the adipose tissue and present in maternal milk, that reduces responses to stress in the infant. We suggest that leptin acts on both central (hypothalamus and hippocampus) and peripheral (pituitary, adrenal gland) targets in the infant to reduce exposure to glucocorticoids and enhance hippocampal development during a sensitive period of brain development. There is also evidence to support the reverse regulatory influence, in which maternal state is profoundly affected by stimulation from the young. During the period of lactation, mothers exhibit lower neuroendocrine and behavioural responses to several types of stressors, except possibly those representing a threat to the infant. This ability to “filter” relevant from irrelevant stimuli while caring for their young might be viewed as adaptive for the mother–infant dyad, and the inability to filter adequately stressful stimuli could at least in part be associated with the development of postpartum depression.
Introduction
Among the complex social relationships observed in altricial mammals, that between mother and infant is possibly the most critical to the provision of optimal conditions for development. Our research efforts have concentrated on the reciprocal nature of this dyad, with respect to the regulation of responsiveness to stress in both mother and offspring (Fig. 1). Beginning in the late 1950s, the pioneering work of Ader and Grota,1 Denenberg et al,2 Levine3 and Levine et al4 established that environmental influences such as handling or infantile stimulation of rats during the neonatal period can lead to long-term changes in both behaviour and response to stress. Over the last 3 decades, extensive efforts have been made to understand how variations in maternal nutritional state or behaviour could influence the development of responsiveness to stress in the infant, potentially leading to long-term consequences for the functioning of the hypothalamic–pituitary–adrenal (HPA) axis. In neonatal rats, for instance, maternal licking and grooming of the pups appears to be particularly effective in maintaining the relative quiescence of the HPA axis during the first 2 weeks of life5–7 and in reducing neuroendocrine and behavioural responsiveness to stress in adulthood.8 In contrast, repeated neonatal maternal separation or exposure of the mother to stress during late gestation or the early postpartum period, or both, have been associated with a number of behavioural impairments and heightened vulnerability to disease in adulthood.9 Early variations in maternal care, whether produced by handling, maternal separation or even naturally occurring, have been associated with molecular changes in the central nervous system (CNS).10–15

Schematic representation of the reciprocal relationships between mother and infant, with particular emphasis on the regulation of responsiveness to stress in both members of the dyad. Knowledge of outcomes directly linked to sensory stimulation and feeding is derived primarily from animal studies. GC = glucocorticoids, HPA = hypothalamic–pituitary–adrenal axis, nesting bout duration = the time the mother spends arched over her pups for them to have access to the nipples.
Although variations in maternal care represent an important factor in the development of the HPA axis (stress axis) and behavioural regulation of the off-spring, maternal state is also profoundly affected by stimulation from the young. Many of the morphological, neurochemical and physiologic changes,16,17 including hyporesponsiveness to a variety of stressors,18–20 that are initiated in late pregnancy in the rat21 are maintained post partum by suckling stimulation from the young. This situation is not unique to rodents. A recent study conducted in breast-feeding women reported decreased plasma adrenocorticotropic hormone (ACTH), cortisol, vasopressin, epinephrine and glucose responses to treadmill stress tests.22
In this paper, we will consider the high fat content of maternal milk and leptin and their effect on the regulation of responses to stress and some aspects of brain development in the rat offspring. We will also examine how the presence of the suckling infant modulates responsiveness to stress in the mother. Finally, the concept of stimulus salience will be discussed in the context of both rodent and human studies.
Mother-to-infant regulation of responsiveness to stress
Developmental aspects of the HPA axis in neonatal rats
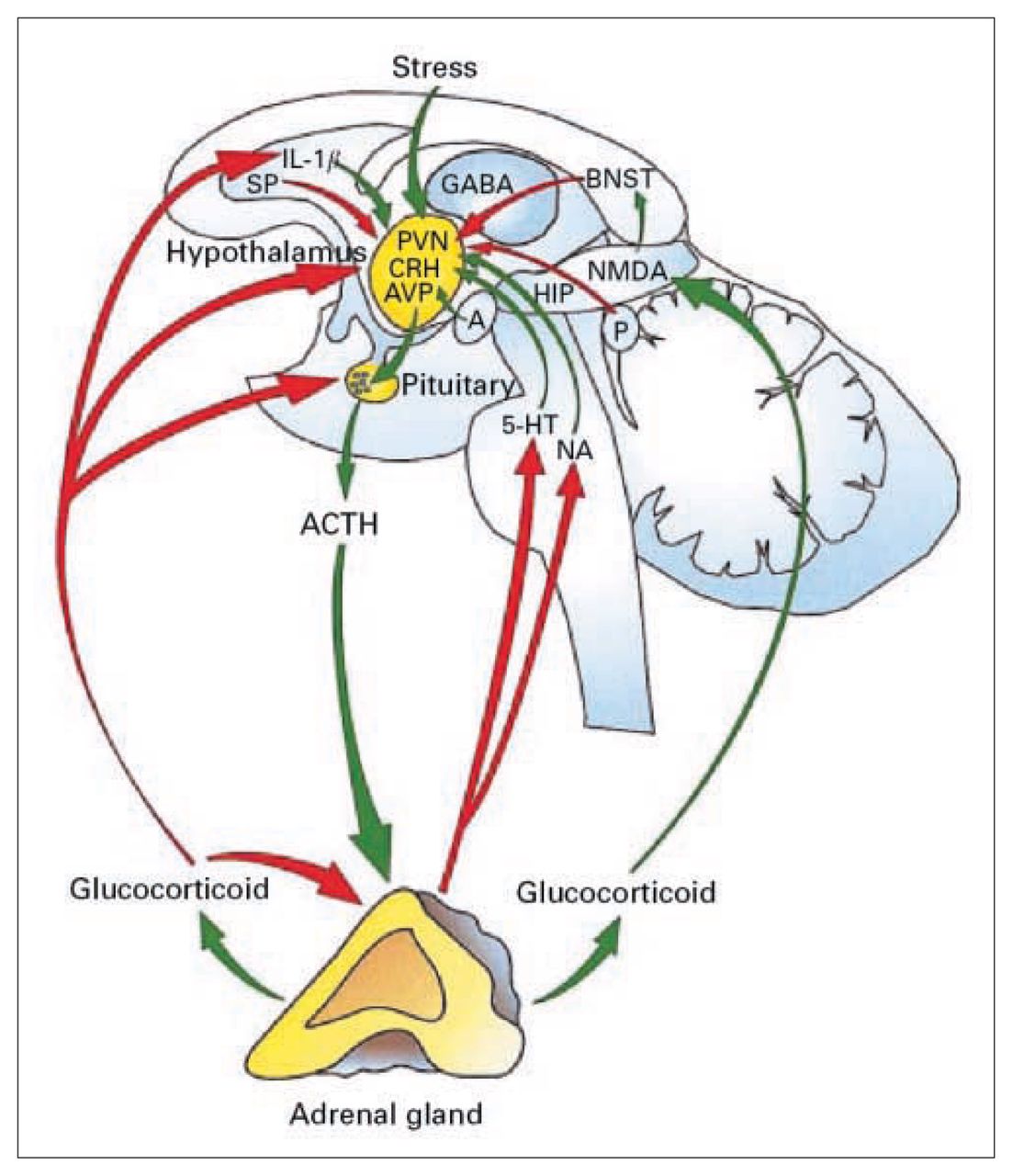
In all mammals, the activation of the HPA axis, which is schematically illustrated in Figure 2,23 is a central component of the integrated physiologic response that occurs after exposure to various internal or environmental stressors, or both. The synthesis and secretion of glucocorticoids that are so critical for developmental processes24,25 represent the final step in a neuroendocrine cascade beginning in the CNS. Somatic and psychologic stressors, circadian drive and humoral influences initiate this cascade by releasing multiple ACTH secretagogues of hypothalamic origin into the hypophysialportal circulation.26–28 Among them, corticotropin-releasing factor (CRF), elaborated by perikarya in the hypothalamic parvocellular paraventricular nucleus (pPVN),29,30 is critical in the regulation of adenohypophysial ACTH secretion27 and proopiomelanocortin (POMC) gene expression.31,32 In rodents, weaker secretagogues, such as arginine vasopressin (AVP), modulate ACTH secretion, but their actions are dependent upon the presence of CRF.33–35 The functional activity of the HPA axis is critically dependent on glucocorticoid feedback mechanisms36,37 that serve to regulate basal levels as well as damp the stressor-induced activation of the HPA axis and to shut off further glucocorticoid secretion.38 Together, these mechanisms permit rapid adjustment of the HPA axis in response to the demands of the environment.

Schematic representation of the activation of the HPA axis in response to stress. Stimulatory pathways to the HPA axis are shown in green, and inhibitory pathways are shown in red. The primary integrator site that receives multiple inputs from the periphery, as well as from central sites, is the hypothalamic paraventricular nucleus (PVN), which contains corticotropin-releasing factor neurons. 1L-1β = interleukin-1β, SP = substance P, GABA = γ-aminobutyric acid, BNST = bed nucleus of the stria terminalis, CRH = corticotropin-releasing hormone, AVP = arginine vasopressin, A = amygdala, P = pineal gland, NMDA = N-methyl-d-aspartate, HIP = hippocampus, 5-HT = serotonin, NA = noradrenaline, ACTH = adrenocorticotropic hormone. Reprinted with permission from Blackwell Publishing Ltd. (J Neuroendocrinol 2001;13:1009–23).23
Information about the internal and external environment reaches the hypothalamic neurosecretory cells over a broad and diffuse neural network.39 Sensory information from the oral cavity, thoracic cavity and the abdominal viscera is carried to the brain stem via the glossopharyngeal and vagus nerves, with other inputs conveyed by the dorsal roots. Cell groups within the nucleus of the solitary tract, A2 and C2 in particular,40 receive baroreceptor41 as well as somatosensory inputs.42 Efferents from the A2, C1, C2 and C3 regions heavily innervate the CRF-rich pPVN area,43 with additional sparse innervation from the ventrolateral medulla (A1) and the locus coeruleus (A6).40,44–46 Emotional stimuli are mediated through corticolimbic pathways that funnel information to the pPVN via the bed nucleus of the stria terminalis, the lateral and medial septum, the fimbria, the amygdala and the preoptic area.47–51 Intrinsic stimuli (e.g., humoral factors, energy substrate availability, core temperature) may be mediated through activity of the vagus nerves, or via humoral agents acting directly on neurosecretory cells or via interneurons.
Following the early work by Selye52 that established criteria for measuring responses to stress in vivo, the pioneering work of Schapiro53 identified a developmental stage in the rat during which responsiveness to stress was abolished; therefore, this period was called “the stress nonresponsive period” (SNRP). However, later studies showed that in marked contrast to stimulated adrenal corticosterone secretion, which is blunted during the first 2 weeks of life in the rat, plasma ACTH responses to a variety of stressors are not significantly altered during this same period.54–57 Qualitative and quantitative plasma ACTH responses to stress in neonates vary as a function of the stressor applied and time after the onset of stress. Stressors such as ether,57,58 cold,54 histamine,54 endotoxin,59 interleukin-1β injection,60 pain,61 social isolation and footshock,62 morphine injection63 or maternal separation5,64–67 result in significant activation of the hypothalamic–pituitary unit. These findings led to a revision of the original concept to that of a “stress hyporesponsive period” (SHRP), which is specifically characterized by blunted adrenal responses to stress during the neonatal period.68 Adrenal stress hyporesponsiveness does not appear to be a simple maturational process, because adrenal responses to stress in the fetal rat in late gestation and in neonates in the first day after birth55,56 are reported to be functionally intact. In contrast to many of the stressors used in neonates, the composite stress of maternal deprivation for 24 hours produces large increases in circulating corticosterone concentrations54,67,69 and increases adrenal sensitivity to ACTH.70,71 Of the 3 main factors that are removed in the maternal separation paradigm (active sensory stimulation, food, passive contact), it has been suggested that milk deprivation may mediate changes in adrenal sensitivity66 and, as will be discussed later, a rapid decline in circulating leptin levels following maternal separation in neonates might constitute a mediating factor. Taken together, these data support the existence of functional hypothalamic–pituitary secretion and effective responses to specific stressors throughout the neonatal period of the rat. Although changes in the time course and pattern of secretion as well as the magnitude of the response or the ratio of POMC products that are secreted can be attributed to maturational processes, evidence is accumulating to support the functionality of the hypothalamic–pituitary unit during the neonatal period in the rat.
The late gestational, perinatal and postnatal periods in most species are characterized by intense synaptic organization in the CNS. This process of synaptogenesis and remodelling underlies the functional maturation of major neurotransmitter and neuromodulatory systems. In later stages of maturation and adulthood, several of these processes can be influenced by glucocorticoid production, the end product of HPA activation. Excess glucocorticoid production is deleterious to brain maturational processes, whereas low adrenal corticosterone secretion in neonatal life allows for adequate brain development. Species differ in their state of CNS maturation at birth. In the rat, the postnatal period is critical for the development and integration of the various central and peripheral systems that maintain homeostasis. In contrast, many of these same systems have reached a greater degree of organization at birth in guinea pigs, sheep, nonhuman primates and humans.
A high-fat diet and leptin regulate stress responses in the offspring
Breast milk is universally agreed to be the single best food for newborns and developing infants in all mammalian species. As noted by Locke,72 “breast milk is a highly complex substance that is more than the sum of its parts.” Bioactive factors such as immunoglobulins, enzymes, growth factors, hormones, glycocongugates and milk lipids can confer immunoprotection and affect development.73 Suckling infants are naturally exposed to a diet that is exceptionally rich in fat, and fat content correlates positively with the length of the interval between periods of suckling across species. Mature human milk from well-nourished communities contains between 3.8 g/100 mL and 4.8 g/100 mL of fat compared with 1.1 g/100 mL of proteins and 6.8 g/100 mL of lactose.74 Although the total fat content and composition of human milk is greatly influenced by the degree of nutrition and the type of diet ingested by the mother, the fact that human milk contains large amounts of long-chain polyunsaturated fatty acids makes this nutrient essential to tissue and organ growth as well as brain development.75 For instance, the accumulation of docosahexaenoic acid (DHA) in brain cortical neurons appears to affect later cognitive development,76 and supplementation of DHA to young infants has been shown to influence the development of visual acuity77 and some aspects of mental development. 77,78 Dietary fat intake and the subsequent changes in brain membrane fatty acid composition have also been implicated in the development of and susceptibility to some mental illnesses.79
Another important component of milk is leptin; this protein, which is produced predominantly by white adipose tissue, can signal the state of energy to the brain and influence food intake and thermogenesis,80–82 as well as a number of other processes related to cellular proliferation and steroidogenesis.83–85 Plasma levels of leptin are naturally high in human neonates86 and developing rodents.87,88 One of the main sources of circulating leptin in neonatal rats is likely to be maternal transfer through milk, because leptin levels rapidly decline to undetectable levels in pups separated from their mothers for 24 hours. An alternative source would be endogenous production by white and brown adipocytes in neonatal rodents.89 In rats, we have demonstrated that leptin content in the milk can be influenced by maternal diet and that feeding a high-fat diet to mothers during the last week of pregnancy and throughout lactation can significantly increase leptin in the milk, as well as circulating levels of leptin in the pups. Conversely, restriction of food to the mother (receiving 60% of the ad libitum ration) during this same period results in extremely low levels of leptin in the offspring. Thus, maternal diet can modify significantly exposure of the offspring to leptin during a critical period of development.
Our interest in leptin arose from earlier studies in which we found that feeding a high-fat diet to the mother could significantly reduce the magnitude of the hormonal (ACTH) response to a stressor in her off-spring. 90 Precedents for the effects of high-fat feeding91 or even a direct effect of fatty acids92 on the activity of the HPA axis were documented in adult animals. Surprisingly, in adult rats, high-fat feeding exacerbates the neuroendocrine response to stress and causes prolonged secretion of glucocorticoids. However, in developing rats, elevated fat in milk appears to blunt the pups’ HPA response, and leptin might be a critical mediator in this effect. Indeed, when pups were administered leptin daily during the first 10 days of life and tested for their ACTH and corticosterone response to stress on postnatal day 10 (PND10), we observed that the overall magnitude of the response was reduced by leptin treatment compared with that in controls. An interesting feature of the response, however, was that the duration of the response was shorter, whereas the peak response was not modified.93 Such a differential pattern of ACTH secretion has been documented in numerous models of chronic stress and aging,94 whereby delayed return to baseline levels of ACTH or corticosterone secretion indicates impairment in the glucocorticoid feedback system. We found that leptin caused a faster return to baseline secretion and, thus, we hypothesized that leptin treatment might have enhanced, rather than impaired, glucocorticoid feedback efficiency on the HPA system.93 To test this hypothesis, we measured the inhibition of stress-induced ACTH secretion by dexamethasone injection in pups that had received long-term treatment with vehicle or leptin at daily doses of 1 mg/kg or 3 mg/kg of birth weight.95 Our results confirmed that long-term leptin treatment can enhance glucocorticoid feedback efficiency in developing rats and showed that part of this effect might be mediated by increases in glucocorticoid receptor expression (mRNA and protein levels) in the hippocampus and hypothalamic PVN, which are both important target sites for glucocorticoid feedback onto the HPA axis.36
Interestingly, a significant relation between leptin and glucocorticoid secretion was also observed in treatment- naive neonates. We found that naturally occurring elevated leptin concentrations in the plasma were inversely related to basal corticosterone secretion during the first 3 weeks of life, which is consistent with results reported in adult rodents and humans.96,97 Because of maternal transfer at the time of delivery, plasma corticosterone levels are high perinatally, decline dramatically over the first few days of life98 and remain low until the third week of life, as described previously. 68 In contrast, plasma leptin concentrations decline briefly at birth and then increase to levels higher than that of the adult rat throughout the suckling period. The robust inverse relation between corticosterone and leptin therefore suggests that leptin might suppress basal adrenal production of corticosterone in developing rat pups. To expand on recent in-vitro studies performed on adult adrenal cells that showed a direct inhibitory effect of leptin on steroidogenesis,99 we conducted both in-vivo and in-vitro studies on freshly dispersed neonatal adrenal cells to address the possible direct effect of leptin on the developing adrenal gland. We tested the response to leptin of adrenocortical cells originating from treatment-naïve pups on PND10 (with normal circulating levels of leptin) and pups separated from their mother for 24 hours, which rapidly reduced endogenous circulating leptin concentrations to undetectable levels. We demonstrated that, in vitro, leptin did not affect basal secretion of corticosterone from adrenocortical cells but significantly reduced the ACTH-stimulated corticosteroid release. The effect of leptin was observed in both treatment-naïve and separated pups, despite the higher basal corticosteroid secretion observed in separated pups.100 Indeed, in these separated pups, the higher level of corticosterone was correlated with a general increase in all the steroidogenic machinery (peripheral benzodiazepine-type receptor [PBR], steroidogenic acute regulatory protein [StAR], 3β-HSD and P450c11).
The effect of leptin was not specific to cells of the fasciculata region of the adrenal cortex that produce corticosterone because, as documented in adults,101,102 ACTH-stimulated aldosterone production from the zona glomerulosa was also reduced by leptin. Thus, it is possible that leptin impaired ACTH receptors in these regions and/or that the inhibitory effect of leptin on adrenal corticosteroid production is attributed to a significant decrease in 2 intracellular proteins (StAR and PBR) that are rate limiting for corticosteroidogenesis. 103–105 Indeed, we found that leptin treatment in vivo reduced the expression of StAR103 and PBR proteins after stimulation with ACTH.106
Hence, in agreement with the findings of earlier studies of the role of feeding in HPA function,70 it is tempting to speculate that naturally occurring high concentrations of leptin in developing pups are critical to maintaining blunted adrenal glucocorticoid secretion during the neonatal period.68 In addition to the regulation of neuroendocrine activity, the functional consequences of opposing actions of leptin and glucocorticoids in rodents on regulation of food intake, body weight gain and thermogenesis have also been investigated in adult rats.107–109 Whereas glucocorticoids appear to limit the action of leptin on food intake in rats108 and on body weight gain and fat stores in normal mice,109 other reports have failed to show a significant interaction between the 2 hormones on food intake and thermogenesis in genetically obese ob/ob mice.107 In developing rodents, the possibility that these 2 hormones have opposing effects on energy balance is difficult to determine using the same parameters, because the primary role of leptin of limiting food intake is not observed in neonatal rats.110
In summary, the maternal regulatory influence on the activity of the HPA axis of the offspring is expressed at least in part through maternal dietary intake of fat contributing to elevated circulating levels of leptin in developing rats. Elevated levels of this hormone might be protective to the brain by maintaining a “brake” on the activity of the HPA axis, and this inhibitory effect can be expressed at various levels of the HPA axis by inhibition of stress-induced CRF mRNA expression,93,111 reduced neuroendocrine responses,93,112 enhanced glucocorticoid feedback efficiency95 and inhibition of adrenal glucocorticoid production.99,100
What are the critical implications of these findings for human nutrition in the postnatal period in term and preterm neonates? As mentioned earlier, leptin is an essential component of maternal human73,113 and cows’114 milk and has recently been found in variable amounts in some infant formula compositions.114 Modulation of leptin levels in milk can be achieved by increased dietary fat intake90 and, in particular, by an increased proportion of n-3 and n-6 polyunsaturated fatty acids,115 as demonstrated in animal studies. However, it is currently not known whether changes in dietary intake in nursing mothers can alter milk leptin levels,73 in addition to changing milk fat content quite dramatically.74,116 Recent studies have demonstrated that serum leptin levels in breast-fed infants are higher than in formula-fed infants during the first year of life,117 although there is no strong relation between milk leptin and adiposity in the infant.118 This does not preclude a more subtle and long-term effect of varying degree of exposure to leptin in the perinatal and early postnatal period in humans. Indeed, leptin might prime or set the endocrine and metabolic system at a different homeostatic level of energy regulation, thus altering the propensity toward obesity.72 There is indirect evidence for this in studies showing that human milk intake is associated with lower leptin concentrations relative to fat mass in adolescence119 and lower risk of being overweight during older childhood and adolescence.120–123 Although leptin is certainly not the only factor responsible for the beneficial effects of breast-feeding on the reduced risk of developing obesity, its relation to fat intake and deposition makes it an important endocrine factor to consider in programming later energy balance and metabolism. In addition to the metabolic regulatory aspect of leptin, this hormone is important for a number of developmental processes including angiogenesis, bone metabolism, hematological differentiation, growth, immune functions and, possibly, brain development as well.124,125 In particular, it has been demonstrated that feeding breast milk, which contains significant amounts of leptin, to premature infants is associated with a reduced risk of gastrointestinal infectious disorders72 compared with that found for formula-fed infants. This suggests that leptin, which is normally supplied by the placenta and the fetus in utero and then supplied through breast milk, could serve as a modulating agent on the immune system of preterm infants.126,127
Putative role of leptin on hippocampal development in pups
Leptin is first and foremost considered to be a hormone that regulates energy balance and food intake in most mammals. In utero, leptin is secreted by the placenta and plays an important role in placental and fetal growth. In the maternal compartment, production of leptin might be important for placental growth, angiogenesis and immunomodulation, whereas leptin produced in the fetal compartment (originating from placenta and fetal adipose tissue primarily) modulates insulin–glucose relations, growth and insulin-like growth factor production as well as immunity.127 The demonstration that leptin levels in the fetal compartment are closely related to birth weight128 emphasizes the critical role of this protein on fetal growth and development. Of leptin’s multiple actions, we have chosen to investigate its putative role in the early development of brain function, because high levels of leptin observed during the perinatal and postnatal periods coincide with a period of intense neurogenesis and synaptogenesis in the CNS.129,130 Typically, circulating plasma levels of leptin are about 5–10 ng/mL in PND10 pups and decrease to 0.5–1.5 ng/mL in weaned rats on PND35 and in adults. The hypothesis that the high leptin levels seen in developing pups play a critical role in brain development and synaptogenesis is based, in part, on the observation that many structural neuronal abnormalities are found in leptin-deficient rodent models (ob/ob mouse, db/db mouse or fa/fa rat).131,132 These abnormalities include reduced brain weight and DNA content, impaired myelinization and increased neurodegeneration in the hippocampus. In the ob/ob mouse, leptin replacement during the postnatal period increases brain growth and DNA content133 and increases synaptic proteins such as syntaxin-1 and synaptosomal associated protein (SNAP-25).134 Evidence in support of this hypothesis comes from our own recent data from DNA microarrays and polymerase chain reaction analysis showing that in normal rat pups, leptin administration increases the expression of several proteins involved in synaptogenesis and neurotransmitter release (synapsin-2A, synaptophysin, SNAP-25) (Table 1). These data are particularly interesting in light of our previous findings that circulating leptin levels increase about 3-fold when pups feed on a high-fat milk, which raises the intriguing possibility that leptin levels are 1 pathway through which early nutritional variables modify brain development. In this context, it is noteworthy that exposure to elevated leptin concentrations coincides with the presence of leptin receptors in the developing brain.135,136 In adults, functional leptin receptors are found in the hypothalamus and extrahypothalamic sites such as the hippocampus, thalamus and cerebral cortex, making these areas susceptible to direct modulation by leptin.135 Among these sites, the developing hippocampus is perhaps the most important for long-term regulation and integration of neuroendocrine stress responses as well as cognitive responses. Extensive neurogenesis and synaptogenesis occurs in this region during postnatal development in rodents and humans,130 and neurogenesis of granule cells in the dentate gyrus persists throughout life, maintaining hippocampal plasticity required for long-term potentiation and memory function.137 In adult rats, leptin has been shown to enhance N-methyl-d-aspartate (NMDA) receptor function via increased Ca2+ influx into hippocampal CA1 neurons. This response to leptin was specific to NMDA receptors, because there were no leptin-induced changes in the electrophysiologic properties of neurons modulated by α-amino-3- hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors.138 Our current work has established that conditions associated with a reduction in circulating leptin levels during the first 2 weeks of life (in pups of food-restricted mothers) results in a significant decrease in NMDA receptor binding in the hippocampus of PND14 pups.139 In addition, daily leptin treatment during the first 2 weeks of life causes long-term changes in the expression of the NMDA receptor subunits NR2A and NR2B mRNA in the hippocampus,140 suggesting that leptin-induced functional modifications in hippocampal activity might persist until adulthood in rodents.
Expression of mRNA levels for selected hippocampal proteins on postnatal day (PND) 10 by DNA membrane microarray*
Although the presence of leptin receptors in the developing hippocampus suggests that leptin may have a direct effect on the development of this structure, leptin could also affect hippocampal function indirectly by increasing glucocorticoid receptor density.95 This effect could be mediated by a reduction in endogenous corticosterone secretion or by accelerated neurogenesis in this particular structure. Hippocampal neurogenesis and synaptogenesis are highly dependent on adequate amounts of glucocorticoids (although not in excess) and glucocorticoid receptors.141 The effect of leptin on hippocampal glucocorticoid receptor expression (in particular in the dentate gyrus) might also be the result of accelerated neurogenesis in this particular structure. We are currently investigating whether leptin acts as a facilitatory factor in neurogenesis and glucocorticoid receptor expression on newly formed neurons (Fig. 3). Whether the actions of circulating leptin on the hippocampus are direct or indirect through modulation of the glucocorticoid environment, the consequences of reduced exposure to this protein during a critical window of brain development cannot be ignored. Similarly, in human fetuses and neonates, reduced placental and/or milk leptin production during reduced intrauterine growth and malnutrition has the potential to impair developmental processes in the CNS. The underlying mechanisms and long-term consequences of inadequate leptin exposure during development on brain functions remain to be elucidated using longterm immunization with antisera or specific receptor antagonists when available.
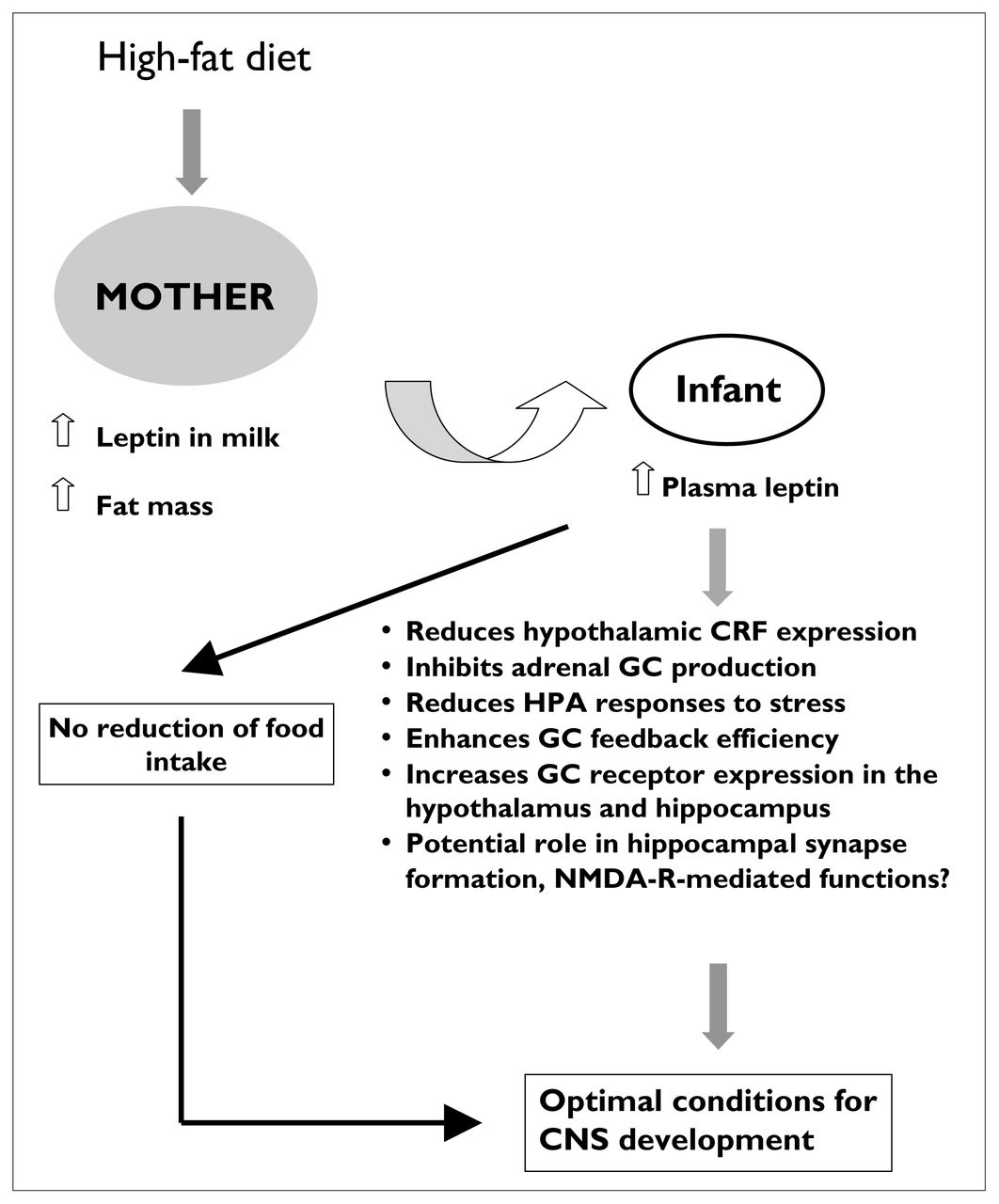
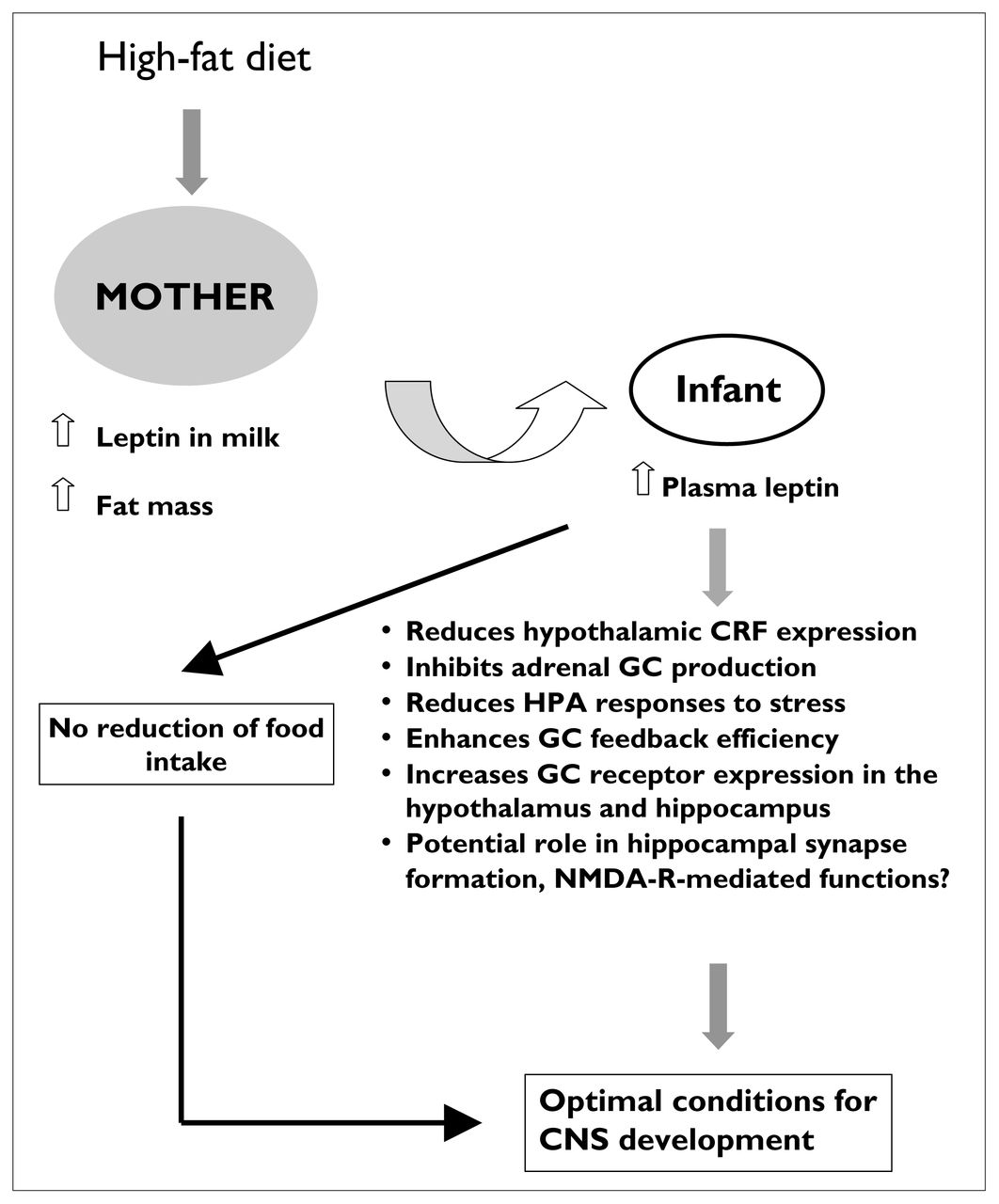
Illustration of the putative mechanisms that mediate the effect on the offspring of high-fat feeding of the mother. Through an increase in maternal fat mass and leptin concentrations in the milk, a maternal high-fat diet increases leptin transfer and circulating levels of this protein in the infant. Despite a lack of effect on food intake in pups on postnatal day 10, leptin has been shown to reduce significantly the activation of the HPA axis after exposure to a stressor and, thus, might be considered to be “protective” during neonatal development. NMDA-R = NMDA receptor, CNS = central nervous system.
Infant-to-mother regulation of responsiveness to stress
There is no question that maternal influence is critical to the optimal physiologic and neurologic development of the offspring. We have noted earlier that maternal nutrition, as well as maternal touch, could represent an important aspect of infant regulation, as described in many other human and animal studies. 8,142,143 The beneficial influences of the mother–infant dyad are certainly not limited to maternal inputs into infant development and extend also in the other direction in ways that have received far less consideration to date. Understandably, much emphasis has been placed on early determinants of adult pathologies in an effort to develop preventive methods and care systems that can adequately address the needs of our present society. It has been known for some time that transgenerational transmission of mothering style can have long-term consequences on the well-being of families,144 and emerging animal studies are starting to unravel some of the putative genetic and/or environmental mechanisms of this transmission.8,145–148 Alternatively, infant temperament might influence mothering style,149–151 and sensory stimuli provided by the infant are known to modify a number of physiologic and behavioural responses in the mother.152–154 In the second part of this paper, we will discuss how the presence of the infant and the suckling stimulus can blunt responsiveness to stress in the mother.
Hyporesponsiveness to stress in nursing mothers
In several species, the period of lactation represents a unique physiologic condition that is well defined in terms of duration, stimulus (suckling) and hormonal milieu and is associated with changes in behaviour,155 metabolic regulation,156 central morphological plasticity16 and neuroendocrine functions.157,158 In particular, female rats during the late gestation period,159,160 parturition161 and lactation display a reduction in HPA-axis responses to a variety of physical and metabolic stressors, including hypertonic saline injection, swim stress, ether stress, lipopolysaccharide injections,19,162,163 as well as psychologic stressors including noise, immobilization or social stress.164–167 This phenomenon is not unique to rodents, because a study conducted in breast-feeding women demonstrated a similar hormonal hyporesponsiveness following the stress of treadmill exercise.22 Several of the hormonal changes of pregnancy and lactation are likely to contribute to the hyporesponsiveness to stress seen at this phase of the life cycle. During the first 2 weeks of lactation, females exhibit high levels of prolactin and oxytocin release,168 tonically elevated corticosterone and progesterone secretion, 163,169 low, diestrus levels of estrogens,170 in addition to decreased central neuronal responsiveness to stimuli.171,172 The state of hyporesponsiveness to stress observed during lactation is thought to result from mechanisms occurring at various sites and spanning the late-pregnancy and lactation periods. These adaptive mechanisms, including modifications to the neuronal phenotype of PVN neurons,17 changes in pituitary sensitivity to ACTH secretagogues160,173 and hormonal variations, are closely linked to the presence of the suckling stimulus, because removal of the pups restores most of the neuroendocrine and morphological changes to normal within 1–3 days. It is worth highlighting at this point that lactation is a very dynamic state both in terms of the distinct regulatory mechanisms being predominant at specific times in the lactation period (early v. late lactation) and the morphological and functional plasticity that is observed during this period in hypothalamic neurons. For instance, potentiated and concerted bursts of oxytocin secretion are possible because of increased synaptic contacts between hypothalamic supraoptic neurons and concomitant glial retraction.16 In our own work, we have demonstrated that the phenotype of hypothalamic PVN neurons changes in lactation to express greater amounts of vasopressin in CRF-expressing neurons compared with those in virgin females.17
Although a detailed investigation of the putative mechanisms that may lead to hyporesponsiveness to stress during lactation is well beyond the scope of this paper, some of these mechanisms deserve particular attention. For instance, lactation induces critical changes in central neurotransmitter release that are both region specific and neurotransmitter specific. Changes in dopaminergic activity have been implicated in prolactin release174,175 and in mediating several aspects of maternal behaviour (e.g., pup retrieval, licking and grooming).176–180 Alterations in serotonergic181 and opioid182,183 activity have also been documented, but the central noradrenergic/adrenergic system, which is a critical regulatory component for HPA activity in normal male and female rats, may be the most studied in relation to the stress or HPA axis.184 The present consensus on the effects of norepinephrine on CRF neurons of the PVN is that α-1 adrenergic receptors mediate the stimulatory effects of norepinephrine at low concentrations (< 5 nmol/L) and that binding to inhibitory presynaptic (and possibly postsynaptic) α-2 and to postsynaptic β-2 adrenergic receptors would be predominant at higher endogenous norepinephrine concentrations and would inhibit the activity of CRF neurons.185 Further inhibition of CRF release would occur through the well-documented action of α-1 adrenergic receptors in potentiating cyclic adenosine monophosphate responses mediated by beta adrenergic receptors.186 Although suckling increases norepinephrine turnover in the rostral PVN and supraoptic nucleus,187 which are sites that contain most of the oxytocinergic neurons, the same suckling stimulus appears to have an opposite effect on PVN neurons that control HPA activity. Indeed, we have previously demonstrated that reduced activity of noradrenergic afferent pathways to the PVN of the hypothalamus and lower secretion of norepinephrine at the level of CRF neurons of the PVN could reduce responsiveness of these neurons to stressors in lactating mothers.188–190 Differential modulation of noradrenergic receptor–mediated activation of PVN neurons160,191 has also been demonstrated. In particular, lactating females have been found to be less responsive to α-1 adrenergic receptor agonists in vivo and on in-vitro electrophysiologic recordings of PVN neurons.191 Taken together, these results suggest that lactation affects different subsets of noradrenergic neurons projecting to the PVN and that reduced responsiveness to stress in lactation might be mediated in part by a reduced ability of noradrenergic terminals to activate pPVN neurons.
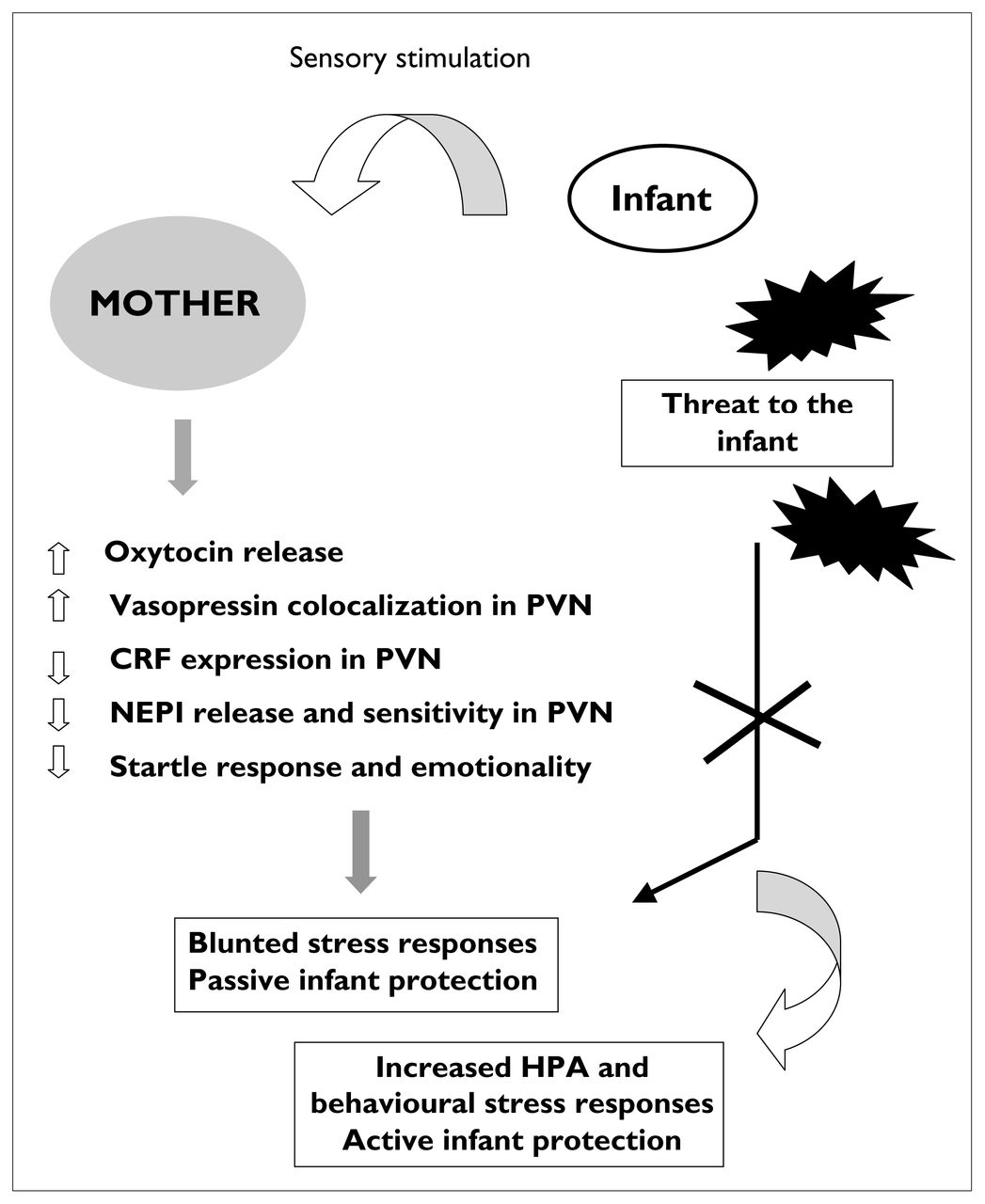
In addition to altering HPA-axis activity and stress responses, there is good evidence that changes in noradrenergic function are also implicated in behavioural regulation and, in particular, emotional responses to acoustic startle. Indeed, lactating females were found to be more sensitive to intracerebroventricular injection of an α-2 adrenoreceptor antagonist, yohimbine, compared with virgin females when tested in the acoustic startle paradigm.192 The altered startle response is only 1 of the multiple behavioural components that are modified in lactation. Lactating females exhibit reduced fear and neophobia165,193,194 and reduced acoustic startle responses,192 and they appear to be less emotional in the open field.167,192 Interestingly, however, the diminished acoustic startle response of lactating females can be quickly restored to normal by fear potentiation, suggesting that the behavioural hyporesponsivity to stress of lactating females might be situation specific. Although we did not measure the neuroendocrine responses to stress in the fear-conditioning paradigm, previous studies have shown that the hormonal response to a stressor in lactating rats can also be modulated by situational variables such as the state of the pups,195 as will be discussed in the next portion of this paper (Fig. 4).

Changes in maternal endocrine and behavioural responsiveness to stressors that are maintained by the sensory stimulation provided by the pups. This schema illustrates our hypothesis that central mechanisms that maintain blunted responses to stress in lactating females can be superseded when the stressor represents a direct threat to the infant. NEPI = norepinephrine.
Pups’ presence modulates responses to stress in mothers
Somatosensory stimulation provided by the suckling pups is critical to maintaining the behavioural, hormonal, morphological and functional changes of lactation. In addition to nipple stimulation at the time of suckling, pups also provide important sensory stimulation to the dam’s ventrum when they are attempting to root to the nipple.196 A comparable type of stimulation has been described in human infants who often massage the breast when they are feeding.152,197 Both types of sensory stimulation might be critical to maintaining neuroendocrine changes in lactating mothers and, in particular, to blunting responsiveness to stress. However, earlier studies have also shown that the magnitude of the maternal response to stress can be influenced by the state of the pups. Mothers reunited with their previously shocked pups showed a greater activation of the HPA axis and, consequently, a greater elevation of plasma corticosterone compared with females reunited with pups that were only handled.195 In this study, it was concluded that the mother responded to specific cues emitted by the shocked pups only when she was not able to have contact with and nurse them. In addition to the state of the pups or what might be better defined as the stimuli associated with particular pup states acting as modulators of the maternal stress response, it is possible that the specific appraisal of the stressful situation by the mother plays an important role in “gating” the neuroendocrine response. This hypothesis is based on our observation that the reduced acoustic startle response of nursing females was rapidly restored to normal after fear potentiation,192 suggesting that the fear component of a stressor could supersede lactation-induced inhibition of the behavioural response to stress. In terms of neuroendocrine regulation, this hypothesis has never been tested, because lactating females are generally exposed to various stressors in the absence of their pups (e.g., ether stress, swim stress, restraint). By analogy, we hypothesized that the magnitude of the neuroendocrine response to stress in lactating females would be dependent upon the saliency of the stressor to the infant and on the ability of the stressor to elicit a fear component. Stressors that represent a direct threat to the infant could provide a sufficient fear component to eliminate hyporesponsiveness to stress and induce potent neuroendocrine and behavioural responses in the lactating mother. We tested this hypothesis by measuring the neuroendocrine responses of lactating females to a psychologic stressor (male intruder or predator odour) in the presence or the absence of their pups. As anticipated, lactating females in the presence of their pups displayed significantly larger responses to psychologic stress compared with the same females tested in the absence of their pups.198 In fact, when tested in the presence of their pups, early lactating females displayed no blunting of the stress response, because the magnitude of the ACTH and corticosterone responses were similar to those in virgin females. This clearly shows that the presence of the pups at the time of exposure to a psychologic stressor enhanced the emotional salience of the stimulus (it changed the stimulus characteristics of the situation) and allowed bypass mechanisms normally in place to blunt the neuroendocrine response. We would like to postulate that specific central mechanisms (e.g., reduced noradrenergic activation of PVN neurons, reduced responsiveness of cortical neurons to NMDA, increased oxytocin and vasopressin expression and function, altered opioid tone) are in place in nursing females to “filter” environmental challenges efficiently and maintain reduced activity in the HPA axis. However, when stimuli that are directly threatening to the pups are presented, these filtering systems are superseded by other mechanisms that allow an adequate neuroendocrine and behavioural response to the stressor. We suspect that part of this activation is provided by structures such as the prefrontal cortex, amygdala and the bed nucleus of the stria terminalis that are implicated in the processing of emotional information as well as the control of the HPA axis. Further work on this circuitry is currently being performed in our laboratory.
As noted previously, lactation is a very dynamic state dictated by varying energetic and behavioural demands placed on the mother by her pups. Our experiments have demonstrated the critical role of the pups’ presence on the regulation of the stress response in the early phase of lactation (i.e., within the first 5 days post partum). During this period, females are behaviourally more responsive to stimuli emitted by pups199 and display greater aggression toward male intruders198,200,201 and larger hormonal responses to stress compared with later in the lactation period.20 Thus, the ability to respond adequately to threatening stimuli might be enhanced compared with later in lactation when the pups are more mobile and less dependent on the presence of their mother for urination/defecation, feeding and thermoregulation. At middle lactation (around PND10–14), central regulatory mechanisms on activation of the HPA axis in the mother might be driven more predominantly by the caloric drain of lactation compared with the appraisal of emotional and threatening situations. Recent elegant data have demonstrated the critical link between caloric intake and suppression of the HPA axis. In these experiments, sucrose ingestion in adrenalectomized rats was as efficient as glucocorticoid replacement at providing inhibitory feedback on hypothalamic CRF neurons.202 It is, therefore, tempting to speculate that the increased food intake observed in middle–late lactating females and the consequent changes in central hypothalamic pathways that regulate metabolism203,204 might lead to major inhibition of the HPA axis that would not necessarily be glucocorticoid dependent. These hypotheses certainly deserve to be tested in the near future.
Implications for human studies
Despite the occurrence of numerous stressors in the early postpartum period (e.g., sleep disturbances, adaptation to a new baby, successful breast-feeding implementation) and the important consequences that postpartum stress might have on the infant, there has been a surprising paucity of studies that have investigated the neuroendocrine responsiveness of postpartum mothers to stress. It is clear that the stress of labour increases cortisol secretion several times in humans205 and animals,18 but only 3 studies to date have measured stress-induced cortisol and autonomic responses during the postpartum and lactation periods in humans.22,206,207 In these studies, it was found that breast-feeding mothers displayed a reduced cortisol and epinephrine response to the physical stress of running on a treadmill22 and that the autonomic response to a psychologic stressor (the Trier Social Stress Test, [TSST]) was blunted compared with that in bottle-feeding mothers.206,207 Interestingly, recent studies conducted in mothers at 1 week post partum208 or 7–8 weeks post partum207 showed that plasma ACTH and cortisol secretion are reduced by infant suckling compared with secretion of these hormones before nursing.208 These studies appeared to compare well with the data reported in the animal literature and support a blunting effect of suckling on the activity of the HPA axis. Our interest, derived from results obtained in our animal studies, was, however, to determine whether stimulus salience could be the basis of the hyporesponsiveness to stress observed in breast-feeding postpartum mothers. We thus reasoned that if a psychologic stressor was relevant to the infant, breast-feeding mothers should display larger responses compared with the response seen with an irrelevant stressor. We used 2 psychologic stressors, the TSST as the irrelevant stressor and viewing of an emotional film suggesting danger to a young child as a salient stressor. In humans, very few studies have measured hormonal responses to the latter experimental protocol. In one study, cortisol and prolactin secretion were significantly enhanced in infertile women watching a video about infertility,209 and their anxiety levels increased throughout the film. In another study, changes in heart rate and blood pressure were found after exposure to an emotional story.210 The experimental conditions that induced emotional reactivity in the laboratory share various peripheral characteristics with real emotions, such as an increase in heart rate, blood pressure and perspiration.211 Also of interest is the observation that reactivity to emotional stimuli in humans has been shown to decrease with exposure and familiarity. 212 For instance, multiparous mothers were found to be less anxious and depressed than primiparous mothers 2 days after delivery.213
The choice of the emotional stressor was also based on the observation that 20%–60% of new mothers experience periods of heightened emotionality or lability, or both, during a postpartum period that can extend over months.214 However, unlike the situation in rodents in which hormones are necessary to maintain the changes of lactation, a host of factors other than hormones contribute to the mother’s mood state post partum.215,216
Although preliminary, our results demonstrate that postpartum mothers still exhibit reduced cortisol responses to psychologic stressors compared with mothers who are not in the postpartum period or with women who are not mothers.217 There was a clear dissociation between ratings of perceived stress and the magnitude of changes in cortisol secretion in all postpartum mothers subjected to the emotional stressor. It is still too early to determine whether stimulus saliency affected the cortisol response or whether there was a significant effect of feeding mode or parity on the neuroendocrine and autonomic responses to the 2 stressors.
In conclusion, nurturing mother–infant interactions are essential for the optimal development of the infant, and numerous behavioural and physiologic regulatory processes ensure the reciprocal nature of the interaction. Regulation of responsiveness to stress in both the mother and infant is essential to achieve adequate behavioural interaction and infant care. Many studies spanning species have demonstrated unequivocally the deleterious effects of disrupted maternal behaviour and infant neglect on cognitive functions and susceptibility to developing disease and drug addiction in the offspring.218 Thus, it appears ecologically sensible that maternal reactivity to stress is selectively blunted toward irrelevant stress and enhanced toward stressors that endanger the mother–infant dyad. It is equally sensible to hypothesize that part of this filtering mechanism is maintained by direct sensory stimulation provided by the infant. Our studies provide some support for the existence of this “filtering mechanism” in rodents and humans; however, we still have many questions that remain unexplored. For instance, we do not know which are the brain structures and the central pathways that support such a “filtering system.” We do not know whether the inability to filter stressful stimuli adequately could at least in part be associated with the development of postpartum depression and whether breast-feeding in high-risk populations could be seen as a protective factor against exaggerated responses to challenges. We hope that answers will be provided to many of these questions in the near future.
Acknowledgements
We would like to acknowledge the expert technical assistance of Geneviè;ve Nault, Kristin Kudreikis, Jessica Jacobczyk and Hong Long in the conduct of these studies. This research has been supported by grants from the Canadian Institutes of Health Research and the Natural Sciences and Engineering Research Council of Canada awarded to Dr. Walker. Dr. Walker is a recipient of a Fonds de la rechereche en santé du Québec senior scholarship award.
Footnotes
Medical subject headings: breast feeding; glucocorticoids; growth and development; human development; lactation; leptin; mice; models, animal; rats; stress.
Claire-Dominique Walker was the recipient of the CCNP Young Investigator Award for 2002.
Competing interests: None declared.
- Received April 9, 2003.
- Revision received July 15, 2003.
- Accepted August 11, 2003.
References

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Article tools






Related Articles
Cited By...
- No citing articles found.