

Abstract
Background: Several current theories emphasize the role of cognitive control in addiction. The present review evaluates neural deficits in the domains of inhibitory control and error processing in individuals with substance dependence and in those showing excessive addiction-like behaviours. The combined evaluation of event-related potential (ERP) and functional magnetic resonance imaging (fMRI) findings in the present review offers unique information on neural deficits in addicted individuals.
Methods: We selected 19 ERP and 22 fMRI studies using stop-signal, go/no-go or Flanker paradigms based on a search of PubMed and Embase.
Results: The most consistent findings in addicted individuals relative to healthy controls were lower N2, error-related negativity and error positivity amplitudes as well as hypoactivation in the anterior cingulate cortex (ACC), inferior frontal gyrus and dorsolateral prefrontal cortex. These neural deficits, however, were not always associated with impaired task performance. With regard to behavioural addictions, some evidence has been found for similar neural deficits; however, studies are scarce and results are not yet conclusive. Differences among the major classes of substances of abuse were identified and involve stronger neural responses to errors in individuals with alcohol dependence versus weaker neural responses to errors in other substance-dependent populations.
Limitations: Task design and analysis techniques vary across studies, thereby reducing comparability among studies and the potential of clinical use of these measures.
Conclusion: Current addiction theories were supported by identifying consistent abnormalities in prefrontal brain function in individuals with addiction. An integrative model is proposed, suggesting that neural deficits in the dorsal ACC may constitute a hallmark neurocognitive deficit underlying addictive behaviours, such as loss of control.
Introduction
The role of cognitive control in substance dependence is emphasized in several contemporary theoretical models.1–6 Individuals with substance dependence are characterized by the inability to adequately inhibit behaviour related to substance use, such as abstaining from substances of abuse. In addition, an apparent failure to adaptively learn from previous harmful behaviour seems characteristic for individuals with substance dependence.7 Inhibitory control and error processing are 2 core components of cognitive control that are associated with specific neural networks: inhibitory control to implement the inhibition of inappropriate behaviour and error processing to monitor performance errors to prevent future mistakes.8 Greater insight into the malfunction of neural networks in individuals with substance dependence underlying inhibitory control and error processing could provide valuable information for understanding the problems associated with controlling substance use. Consequently, a rapidly increasing number of studies have examined inhibitory control and error processing in individuals with substance dependence by using neuroimaging techniques, such as event-related potentials (ERPs) and functional magnetic resonance imaging (fMRI). A combined review of ERP and fMRI studies may provide valuable and complementary insights on both temporal and spatial properties of the neural substrate of problems associated with inhibitory control and error processing in individuals with substance dependence. Therefore, the main aim of the present review is to evaluate the consistency of findings of fMRI and ERP studies investigating inhibitory control and error processing in the major classes of substance-dependent populations.
A second goal of this review is to contribute to the ongoing discussion about the differences and similarities between substance dependence and other excessive behaviours that have been proposed to be related to addiction but that do not involve the ingestion of substances.9 For example, pathological gambling is characterized by unsuccessful efforts to control, cut back or stop gambling, similar to problems controlling substance use. Based on these and other similarities,10–12 pathological gambling is listed under the heading “substance use and addictive disorders” in DSM-5. Other suggested behavioural addictions, such as excessive eating,13 computer game playing or Internet use9 are not included as behavioural addictions in DSM-5 because of a current lack of sufficient scientific evidence for similar dysfunctions in people with these behaviours and those with substance dependence. To contribute to this ongoing discussion and to identify possible gaps in the literature, we systematically reviewed neuroimaging studies that investigated inhibitory control and error processing in people with pathological gambling and those with excessive eating, gaming or Internet use. Throughout this paper, the term “addiction” refers to both substance dependence and the proposed behavioural addictions.
This review starts with an explanation of the experimental task paradigms most frequently used to measure inhibitory control and error processing. In addition, neural correlates of inhibitory control and error processing are discussed to provide a framework for the evaluation of empirical studies. The literature review is organized according to primary substance of abuse (i.e., nicotine, alcohol, cannabis, stimulants and opioids), with a separate section for excessive addiction-like behaviours. This review will conclude with a discussion of the findings, including an integrative model of the findings and future research directions.
Experimental measures and neural correlates of inhibitory control and error processing
Inhibitory control
Experimental measures of inhibitory control
The go/no-go and stop-signal tasks are most commonly used to measure inhibitory control.14–16 In the go/no-go task, participants respond as quickly as possible to frequent go stimuli and inhibit responses to infrequent no-go stimuli, which requires inhibitory control to overcome automatic response tendencies. The proportion of correctly inhibited no-go trials reflects the ability to inhibit automatic behaviour. The stop-signal paradigm17 measures the ability to exert inhibitory control over a response that has already been initiated by asking participants to respond as quickly as possible to a continuous stream of go stimuli. In a minority of the trials, a stop signal is presented after the onset of the primary stimulus indicating that the response to this stimulus should be cancelled. The ability to inhibit already initiated behaviour is indexed by the stop-signal reaction time (SSRT), which is the time needed to cancel 50% of the stop trials relative to mean reaction time for go stimuli. Larger SSRTs represent worse inhibitory control. Most stop-signal paradigms use a staircase method, implying that the number of errors in the task is deliberately kept constant to calculate the SSRT. Although we believe that both the go/no-go and the stop-signal tasks require the activation of a common inhibitory brake, we are also aware that more general processes, such as attentional monitoring and salience processing, may play a role in these tasks.18–20 Besides the go/no-go and the stop-signal tasks, other cognitive paradigms, such as the Stroop21 and Eriksen Flanker22 tasks have been argued to measure inhibitory capacities. However, these tasks also measure other processes, such as conflict resolution, response selection and attention.23,24 To keep the present review focused and to be able to make straightforward comparisons of results, we included only studies using go/no-go and stop-signal paradigms.
Event-related potential measures of inhibitory control
Two ERP components have been reported to reflect changes in brain activity related to inhibitory control.25 The first component, the N2, is a negative-going wave emerging 200–300 ms after stimulus presentation. The neural generators of the N2 appear include the anterior cingulate cortex (ACC)25–27 and the right inferior frontal gyrus (IFG).28 The N2 is believed to index a top–down mechanism needed to inhibit the automatic tendency to respond29,30 and corresponds to behavioural outcomes of inhibitory control.31–33 The N2 has further been associated with conflict detection during early stages of the inhibition process.27,29 Consequently, the N2 can be interpreted as an index for early cognitive processes necessary to implement inhibitory control rather than the actual inhibitory brake. The P3, the second ERP component involved in inhibitory control, is a positive-going wave emerging 300–500 ms after stimulus onset. The source of the P3 has been found to be close to motor and premotor cortices.25,26,34 Hence, P3 amplitudes appear to reflect a later stage of the inhibitory process closely related to the actual inhibition of the motor system in the premotor cortex.25,33,35 Together, accumulating evidence suggests that the N2 and P3 reflect functionally distinct processes associated with inhibitory control. Accordingly, less pronounced N2 or P3 amplitudes in addicted populations relative to controls can be considered markers for neural deficits in inhibitory control.
Functional MRI measures of inhibitory control
Inhibitory control in healthy individuals is associated with a mainly right lateralized network, including the IFG, ACC/pre–supplementary motor area (SMA) and dorsolateral prefrontal cortex (DLPFC) as well as parietal and subcortical areas, including the thalamus and basal ganglia.15,36,37 Experimental studies have provided information on the specific contribution of these regions in implementing inhibitory control. A recent hypothesis suggests that the right IFG, in inhibitory control, detects behaviourally relevant stimuli (e.g., no-go or stop-signal stimuli) in cooperation with the inferior parietal lobe (IPL) and temporal parietal junction (TPJ) through its effects on stimulus-driven attention, which is a crucial element of both go/no-go and stop-signal task performance.18–20 Given the proximity of the pre-SMA/dorsal ACC (dACC) to the motor areas, the function of this region may be response selection and updating motor plans.38 In addition to frontal and parietal regions, the involvement of subcortical regions in inhibitory control is well established through feedback loops that connect these regions with pre-frontal and motor areas.15,36,39 As an extensive basis of fMRI studies has consistently shown that activation in this cortical–striatal–thalamic network is linked to inhibitory control in healthy participants, differences in brain activation in this network during the performance of inhibitory control paradigms in individuals with addictions relative to controls can be interpreted as the presence of neural deficits in inhibitory control in these individuals.
Error processing
Experimental measures of error processing
The most commonly used paradigms are the Eriksen Flanker and the go/no-go task.40,41 In a typical version of the Flanker task, participants are exposed to series of letters. In the congruent condition, 5 equal letters are presented, while in the incongruent condition the middle letter differs from the other letters (e.g, SSHSS/HHSHH). Participants are asked to identify the middle letter. The high stimulus conflict situation in the incongruent condition usually results in performance errors. False-positive errors observed in go/no-go or stop-signal paradigms, are also used to evaluate error-processing. Regardless of the task paradigm, reaction times on trials after performance errors are usually longer than reaction times on trials following correct responses, a process referred to as post-error slowing. Reaction times, the number of errors and this post-error slowing are all regarded as behavioural indices of error monitoring.42,43
Event-related potential measures of error processing
Event-related potential investigations of error processing have revealed 2 error-related brain waves that consistently emerge after performance errors (i.e., error-related negativity [ERN] and error positivity [Pe]). The ERN and Pe appear to be independent as they are differentially sensitive to experimental manipulations and individual differences in task performance, and they reflect different stages of error processing.40,44,45 The ERN arises 50–80 ms after making an error and is known to reflect initial and automatic error detection.46 Converging evidence indicates that the ACC is the neural generator of the ERN.8,47–50 The ERN is followed by the Pe, a positive deflection observed on the electroencephalogram (EEG), emerging approximately 300 ms after incorrect responses.51 Research identifying the neural origin of the Pe has provided heterogeneous results.52 Conceptually, the Pe appears to be associated with the more conscious evaluation of errors, error awareness,40,52 and with the motivational significance attributed to an error.53 Together, the ERN and Pe evaluate the correctness of ongoing behaviour (i.e., a specific outcome or behaviour was worse or better than expected), which is used to guide future behaviour54 and can be used as a neural marker of error processing in individuals with addictions.
Functional MRI measures of error processing
The crucial role for the ACC in error processing suggested by ERP studies has been confirmed in fMRI studies. More specifically, Ridderinkhof and colleagues24 suggest that the dACC/pre-SMA, is consistently activated during monitoring of ongoing behaviour. Some researchers suggest that this region monitors response conflict or the likelihood of errors55,56 rather than error processing per se. Two independent meta-analyses have shown that both response conflict and response error activate the dACC.8,57 Functional MRI studies investigating error processing further show that a large neural network coactivates with the dACC, including the bilateral insula, DLPFC, thalamus and right IPL.57,58 Functional interactions among these regions have been reported, especially between the dACC and the DLPFC.59 Performance errors in the human brain are processed by a neural circuit that extends beyond the dACC and includes the insula, DLPFC, thalamus and parietal regions. This error processing circuit collectively monitors and adjusts behaviour when necessary. As the neuroanatomical substrate of error processing has consistently been demonstrated in fMRI studies in healthy participants, activation differences between individuals with addictions and controls in this error processing network may be interpreted as a neural correlate of possible error-related deficits in individuals with addictions.
Literature review
Selection of studies
We performed a literature search on PubMed and Embase using search medical subject headings (MeSH) for substance-dependent populations and populations with possible behavioural addictions. The MeSH terms were “substance-related disorders,” “alcohol-related disorders,” “amphetmine-related disorders,” “cocaine-related disorders,” “marijuana abuse,” “opioid-related disorders,” “gambling,” “obesity,” “bulimia” and “eating disorders.” We also searched using the keywords “smokers,” “gaming,” “gamers” and “Internet.” The key search terms for various addicted populations had to co-occur in combination with the following search terms concerning inhibitory control and error processing: “cognitive control,” “inhibitory control,” “response inhibition,” “error processing,” “error monitoring,” “go/no-go,” “stop-signal” or “Flanker.” They also had to co-occur in combination with the following search terms for neuroimaging measures: “magnetic resonance imaging,” “evoked potentials” (MeSH terms), “error-related negativity,” “error positivity,” “N200,” “N2,” “P300” and “P3.” The search was limited to research performed in humans and articles written in English. All included articles were required to be published in peer-reviewed journals and indexed in PubMed or Embase before June 2013.
We screened a total of 207 abstracts for the following inclusion criteria: inclusion of a group of individuals with addictions or individuals showing behavioural addictions (social drinkers and recreational drug users were not included); inclusion of a control group such that hypoactivation or hyper-activation as well as behavioural deficits described in this review are always relative to healthy controls (studies without a control group were included only if they evaluated the effect of a treatment outcome or a pharmacological intervention within the addiction group); inclusion of more than 10 participants in each group; us of the go/no-go, stop-signal or Eriksen Flanker task as a measure for inhibitory control or error processing; and the use of fMRI or ERPs as neuroimaging tools. A total of 36 studies fulfilled our inclusion criteria. We manually searched the references in those 36 articles, which yielded another 5 studies that met our inclusion criteria. In all, we included 41 studies in our review. Table 1, displays all relevant participant characteristics, such as age, sex, abstinence, disorder and treatment status. Results of all studies are summarized in Tables 2 and 3, and are discussed in the sections that follow. We refer to the tables for study details, such as participant characteristics and within-subject contrasts, that were used for between-subject analyses in our discussion of these results.
Patient characteristics of included studies
Overview of ERP and fMRI studies investigating inhibitory control in substance dependence and behavioural addictions (part 1 of 3)
Overview of ERP and fMRI studies investigating error processing in substance dependence and behavioural addictions
Inhibitory control
Inhibitory control in individuals with nicotine dependence
We identified 2 ERP studies in the domain of inhibitory control in individuals with nicotine dependence. Evans and colleagues60 investigated inhibitory control in participants with nicotine dependence (abstinence 0–10.5 h) and controls by evaluating P3 (but not N2) amplitudes in a go/no-go task. While no-go P3 amplitudes were lower in those with nicotine dependence than controls, no performance differences between the groups were found. Luijten and colleagues61 investigated whether inhibitory control in nicotine-dependent individuals who had abstained from smoking for 1 hour was influenced by the presence of smoking cues. Compared with controls, those with nicotine dependence were less accurate on no-go tasks and exhibited lower no-go N2 amplitudes. The P3 amplitudes did not differ between groups. Interestingly, behavioural deficits as well as lower N2 amplitudes in individuals with nicotine dependence were found during exposure to both smoking-related and neutral pictures, suggesting that the observed deficit in inhibitory control reflects a general inhibition problem that is not further impaired when smoking cues are present.
We also included 5 fMRI studies of inhibitory control in smokers. One of the key regions involved in inhibitory control, the dACC, was less active in individuals with nicotine dependence than controls during performance of the stop-signal task, while SSRTs did not differ.62 Using a go/no-go task, Nestor and colleagues63 found behavioural deficits for inhibitory control in nonabstinent individuals with nicotine dependence compared with both healthy controls and ex-smokers who were smoke-free for at least 1 year. In addition, the finding of lower brain activation associated with inhibitory control in those with nicotine dependence compared with controls in the ACC was confirmed in this study and ex-tended to the right superior frontal gyrus (SFG), left middle frontal gyrus (MFG), bilateral IPL and middle temporal gyrus (MTG). The nicotine-dependent and ex-smokers groups both showed less activation in the left IFG, bilateral insula, paracentral gyrus, right MTG and left parahippocampal gyrus (PHG) than controls. These results suggest that behavioural and activation deficits in individuals with nicotine dependence may be reversible to some extent, while hypo-activation in other regions persists even after prolonged periods of abstinence. An alternative interpretation may be that in heavily dependent smokers there is an association between the more pronounced behavioural and neural deficits and the failure to give up smoking. The findings of a study involving adolescents with nicotine dependence who abstained from smoking for 30–1050 minutes before scanning support this hypothesis.64 While adolescents with nicotine dependence and controls had similar accuracy rates and brain activation, the study found that severity of smoking within those with nicotine dependence was associated with lower activation in regions critically involved in inhibitory control (i.e., ACC, SMA, left IFG, left orbitofrontal cortex [OFC], bilateral MFG and right SFG).
The pharmacology of inhibitory control in individuals with nicotine dependence and controls was investigated in an fMRI study using a double-blind randomized crossover design with placebo and the dopamine antagonist haloperidol.65 The nicotine-dependent individuals did not smoke for at least 4 hours before the go/no-go task performance. Behavioural findings showed lower no-go accuracy during the first test as well as hypoactivation in the right ACC and MFG and the left IFG after placebo in individuals with nicotine dependence compared with controls. Hyperactivation in participants with nicotine dependence after placebo was found in the right TPJ, which may constitute an attentional compensation mechanism.18 After administration of haloperidol, hypoactivation in those with nicotine dependence relative to controls was found only in the right ACC but no longer in the right MFG and left IFG. Activation patterns suggest that similar brain activation for individuals with nicotine dependence and controls after administration of haloperidol is most likely due to a reduction in brain activation in controls caused by haloperidol. These findings suggest that reduced dopaminergic neurotransmission may be disadvantageous for inhibitory control, which was further supported by the findings that no-go accuracy rates as well as brain activation in the inhibitory control network (i.e., the left ACC, right SFG, left IFG, left posterior cingulate gyrus [PCC] and MTG) were reduced across groups after haloperidol administration compared with placebo. These findings provide valuable information regarding the role of dopaminergic neurotransmission on inhibitory control and suggest that altered baseline dopamine levels in individuals with addictions may contribute to problems with inhibitory control in these individuals.
Berkman and colleagues66 investigated the link between brain activation during inhibitory control on a go/no-go task and real world inhibition of craving. Individuals with nicotine dependence reported cravings and number of smoked cigarettes several times during the first 3 weeks after a quit attempt. The study found that higher brain activation associated with inhibitory control in the bilateral IFG, SMA, putamen and left caudate attenuated the association between craving and real world smoking, while an association in the opposite direction was found for the amygdala. Two important conclusions can be drawn from this study. First, brain activation in an abstract laboratory task to measure inhibitory control is associated with inhibition of feelings of craving in daily life. Second, lower brain activation in regions critical for inhibitory control is actually disadvantageous because it is associated with a strong coupling between craving and smoking.
Summary
The 2 ERP studies provide tentative evidence that N2 amplitudes may be lower in individuals with nicotine dependence than controls, while results for P3 amplitudes are contradictory. Functional MRI studies show hypoactivation in the inhibitory neural network that may be associated with severity of smoking and could be partly reversible after smoking cessation. Hypoactivation during inhibitory control has been shown to be disadvantageous for smoking behaviour, as it was associated with increased coupling between craving and smoking after a quit attempt. Notably, hypoactivation associated with inhibitory control in individuals with nicotine dependence was not always accompanied by behavioural deficits, thereby complicating the interpretation of some of the observed findings. Furthermore, dopaminergic modulation appears to influence inhibitory control capacities.
Inhibitory control in individuals with alcohol dependence
All studies included in this section involve abstaining individuals with alcohol dependence who were currently enrolled in treatment programs. We identified 7 ERP studies for inclusion in this section, 6 of which evaluated P3 amplitudes related to inhibitory control. Kamarajan and colleagues67 found that individuals with alcohol dependence were less accurate than controls during task performance, whereas the other studies did not observe accuracy differences between individuals with alcohol dependence and controls. In 3 studies, smaller no-go P3 amplitudes were observed in individuals with alcohol dependence compared with controls.67–69 However, some of these and other studies also found less pronounced P3 amplitudes for go trials,67,68,70 suggesting that group differences in these studies do not merely reflect differences in inhibitory capacities but rather may be related to more general deficits (e.g., attention). In contrast, Karch and colleagues71 and Fallgatter and colleagues72 did not find deficits in individuals with alcohol dependence on either go or no-go P3 amplitudes. Comparison of these studies is hampered by considerable methodological differences. First, task paradigms differed greatly among studies: in some studies go and no-go probabilities varied across blocks70 or no-go probabilities were high, resulting in low inhibitory requirements.67,72 In addition, some task paradigms involved reward evaluation67 or cueing for no-go trials.72 Second, data analyses in some studies were not focused on regions in which no-go amplitudes usually peak68 or were focused on P3 localization rather than amplitudes.72 Altogether, evidence for neural deficits in the later stages of inhibitory control in individuals with alcohol dependence is mixed, most likely as a result of large methodological differences. One of the included ERP studies investigated N2 amplitudes in participants with alcohol dependence.73 In this study, no behavioural deficits were found for no-go accuracy, while participants with alcohol dependence were less accurate on go trials and showed lower go and no-go N2 amplitudes compared with controls.
We identified 3 fMRI studies for inclusion in this section. Notably, as brain activation was simultaneously measured with EEG and fMRI, the fMRI study by Karch and colleagues74 involves the same patients as the described ERP study by the same group.71 The fMRI findings in these patients confirm ERP findings of comparable brain activation levels for individuals with alcohol dependence and controls.74 The fMRI studies using the stop-signal task in participants with alcohol dependence and controls did not show group differences in SSRTs.75,76 Nevertheless, lower activation patterns associated with inhibitory control in the left DLPFC in those with alcohol dependence could be demonstrated.75 In a pharmacological intervention study, effects of a single dose of the cognitive enhancer drug modafinil on response inhibition and underlying neural correlates were investigated in a randomized, double-blind, placebo-controlled crossover study.76 No main effect of modafinil on SSRT was observed. However, a positive correlation between SSRT after placebo and improvement in SSRT after modafinil suggests that participants with lower baseline inhibitory control may benefit from modafinil. The change in SSRT in individuals with alcohol dependence after modafinil administration was associated with increased activation in the left SMA and right ventrolateral thalamus, suggesting that this may be the neural correlate of improved inhibitory control after modafinil administration in patients with poor baseline inhibitory control.
Summary
As only 1 study evaluated N2 amplitudes, no firm conclusions can be formulated regarding early inhibitory control processes in individuals with alcohol dependence. Evidence for neural deficits on P3 amplitudes reflecting inhibitory control in these individuals is weak, most likely owing to large methodological differences among studies and general study limitations. Some findings in the studies we reviewed suggest that P3 deficits in individuals with alcohol dependence during inhibition-related task performance may be due to general cognitive deficits, such as attention. Specific behavioural deficits for inhibitory control were not convincingly shown in either ERP or fMRI studies, which is in line with conflicting findings in behavioural studies in this domain.77–80 While the number of fMRI studies is limited, available fMRI results suggest that activation in the DLPFC related to inhibitory control in individuals with alcohol dependence may be dysfunctional. Furthermore, inhibitory control in patients with poor baseline inhibitory control may be improved with the cognitive enhancer modafinil.
Inhibitory control in individuals with cannabis dependence
Currently, no published ERP studies involving individuals with cannabis dependence have evaluated N2 or P3 amplitudes in the context of inhibitory control, whereas 2 fMRI studies have been published.81,82 Neither fMRI study found inhibitory control deficits in individuals with cannabis dependence (using go/no-go tasks), which is line with the results of nonimaging studies in similar populations.83,84 However, individuals actively using cannabis showed increased activation during inhibitory control relative to controls in the ACC/pre-SMA, right IPL and putamen.81 These findings can be interpreted as a compensatory neural mechanism, given that individuals with cannabis dependence did not show behavioural deficits. A similar result was also found in abstaining adolescents with cannabis dependence, who showed increased activation during inhibitory control relative to controls in a large network of brain regions (Table 2).82 However, activation in part of these regions was also higher in those with cannabis dependence than controls during go trials, suggesting that not all differences between groups were specific for inhibitory control.
Summary
Clearly, more research is needed to confirm initial fMRI findings that individuals with cannabis dependence need greater neural activation in prefrontal and parietal regions to perform inhibition tasks at the same level as controls. In addition, the time course of possible neural deficits in individuals with cannabis dependence should be investigated by measuring N2 and P3 amplitudes.
Inhibitory control in individuals with stimulant dependence
In 1 ERP study, N2 and P3 amplitudes were evaluated in a Flanker task that incorporated no-go trials in currently using individuals with cocaine dependence.85 The study found that enhancement of no-go N2 and P3 amplitudes relative to go amplitudes was less pronounced in individuals with cocaine dependence than controls. However, behavioural findings did not show differences in accuracy, such that ERP results should be interpreted cautiously.
We included 6 fMRI studies in this section, 5 of which involved patients with cocaine dependence and 1 involved patients with methamphetamine dependence. The studies of Hester and Garavan86 and Kaufman and colleagues87 both found lower no-go accuracy in individuals currently using cocaine accompanied by reduced activation in the ACC/pre-SMA compared with controls. Less brain activation associated with inhibitory control in those with cocaine dependence relative to controls was found in the right superior frontal gyrus86 and right insula.87 The go/no-go task in the study by Hester and Garavan86 involved different levels of working memory load in an attempt to mimic the high working memory demands resulting from drug-related ruminations. The hypoactivation associated with inhibitory control in the ACC was most pronounced when working memory load was high, suggesting that inhibitory control is most compromised in situations requiring high working memory demands. Using a stop-signal task, Li and colleagues88 confirmed hypoactivation associated with inhibitory control in the ACC in abstaining individuals with cocaine dependence relative to controls; this hypoactivation was extended to the bilateral superior parietal lobe (SPL) and left inferior occipital gyrus. However, no differences were found between groups regarding behavioural measures reflecting inhibitory control (SSRTs), which is in contrast to findings from the studies using go/no-go tasks in active users. No association between inhibitory control–related brain activation and relapse rates after 3 months was found in a study of abstaining individuals with cocaine dependence.89
Two fMRI studies involving patients with stimulant dependence investigated possible strategies to improve inhibitory control. A pharmacological fMRI study in abstaining patients with cocaine dependence90 showed that methylphenidate administration enhanced inhibitory control in these individuals (i.e., the SSRT was shorter after methylphenidate administration). Furthermore, methylphenidate-induced decreases in SSRT were positively correlated with activation in the left MGF and negatively correlated with activation in the right ventromedial prefrontal cortex, suggesting that these regions may constitute a biomarker for the methylphenidate-induced increase in inhibitory control. Generally, methylphenidate increased brain activation during inhibitory control in the bilateral striatum, bilateral thalamus and right cerebellum and decreased activation in the right superior temporal gyrus (STG). These differences in activation may also indirectly contribute to the improvement in inhibitory control due to methylphenidate. Another study in abstaining individuals with methamphetamine dependence that used a go/no-go task, did not find evidence for impaired performance or brain activation associated with inhibitory control in these individuals.91 Nevertheless, the study found that accuracy for no-go trials was enhanced in individuals with methamphetamine dependence (and not in controls) when no-go trials were preceded by an explicit warning cue that signalled the need for inhibition on the next trial. In addition, individuals with methamphetamine dependence showed increased activation in the ACC for warning cues, which was positively correlated with improved accuracy. These findings imply that inhibitory control can be improved by explicit environmental cues that predict the need for inhibitory control via preactivation of the ACC. Alternatively, individuals with methamphetamine dependence may benefit from exogenous cues by boosting attention to no-go stimuli. However, a first attempt to link inhibitory control–related brain activation with relapse did not identify brain regions that differentiated between patients who relapsed and those who remained abstinent.89
Summary
Several conclusions can be drawn from the neuroimaging studies in individuals with stimulant dependence. First, the single ERP study in those with cocaine dependence suggests that neural deficits may be present in both early and late stages of the inhibition process; however, it is unclear whether this may result in behavioural deficits. Second, hypoactivation in the ACC during inhibitory control in individuals with cocaine dependence was found, which was associated with impaired task performance in 2 studies. Third, explicit external cues and methylphenidate may both improve inhibitory control by increasing activation associated with inhibitory control in the medial prefrontal cortex.
Inhibitory control in individuals with opiate dependence
So far, 1 ERP study has investigated inhibitory control in abstaining individuals with opiate dependence in which no differences between groups on no-go accuracy or N2 and P3 amplitudes were found.92 It should be noted, however, that inhibitory requirements in this task were low given the high probability of no-go trials (i.e., 50% of the trials were no-go trials), so that the task may have been too easy to reveal differences in inhibitory control between those with opiate dependence and controls.
The single fMRI study included in this section used a go/no-go task in which accuracy levels were deliberately kept constant across individuals. Abstaining individuals with opiate dependence were found to have slower go reaction times and less brain activation than controls during task performance in the key regions implicated in inhibitory control, such as the bilateral ACC, medial PFC, bilateral IFG, left MFG, left insula and right SPL.93 Hypoactivation in individuals with opiate dependence was also extended to regions outside the inhibitory control network into the left uncus, left PHG, right precuneus and right MTG. However, go and no-go stimuli in this study were presented in blocks, such that inhibitory requirements were very low.
Summary
The single ERP study that we included did not show deficits in inhibitory control and associated ERPs in abstaining patients with opiate dependence, whereas hypoactivation in medial, dorsolateral and parietal regions was found in the fMRI study. Generally, studies investigating inhibitory control in individuals with opiate dependence are scarce and, as inhibitory requirements were low in both reviewed studies, future studies could benefit from improvements in task design.
Inhibitory control in individuals with behavioural addictions
We included 3 ERP studies investigating inhibitory control in people with behavioural addictions, 2 of which studied excessive Internet use and 1 of which studied excessive gaming. The ERP study by Zhou and colleagues94 showed less pronounced no-go N2 amplitudes and lower no-go accuracy in excessive compared with casual Internet users. The study did not evaluate P3 amplitudes. Dong and colleagues95 confirmed less pronounced no-go N2 amplitudes in men with excessive Internet use than in those with casual Internet use, whereas P3 amplitudes in those with excessive Internet use were enhanced. No differences in behavioural performance were found in the latter study. Enhanced activation in the final stage of inhibitory control could have served as a compensation for the less efficient early inhibitory mechanisms in excessive Internet users to obtain behavioural performance levels equal to those of casual Internet users. Findings in a third ERP study96 confirm problems with inhibitory control in individuals with behavioural addictions, as excessive gaming in this study was found to be associated with lower no-go accuracy. The ERP findings, however, contradict those of the other studies by showing larger no-go N2 amplitudes in excessive gamers in a parietal cluster compared with controls. Inconsistencies in N2 findings may be the result of differences in study population (a mixed group of excessive Internet users versus a group with only excessive gaming behaviour) or differences in task difficulty (> 91% no-go accuracy across groups in the studies by Dong and colleagues95 and Zhou and colleagues94 v. 53% in the study by Littel and colleagues96).
We included 4 fMRI studies in this section, 2 of which involved individuals with pathological gambling and 2 of which involved participants with excessive eating behaviours. One of the fMRI studies of individuals with pathological gambling reduced activation in the dACC for successful stops in a stop-signal task relative to controls.62 Although SSRTs were not impaired in the pathological gambling group, this finding suggests hypoactivation in the dACC similar to that found in individuals with substance dependence. Another study of individuals with pathological gambling that used a go/no-go task with neutral, gambling, positive and negative pictures showed similar no-go accuracy rates for the pathological gambling and control groups.97 However, those with pathological gambling may have used a compensation strategy to perform the task as accurately as controls, as go reaction times were longer and brain activation associated with neutral inhibitory control in the bilateral DLPFC and right ACC was higher in the pathological gambling group than the control group. A gambling-related context appears to facilitate response inhibition in individuals with pathological gambling relative to controls, as indicated by higher no-go accuracy during exposure to gambling cues and lower brain activity in the DLPFC and ACC in those with pathological gambling than controls.
Two fMRI studies investigating inhibitory control have been performed in people with excessive eating behaviour (i.e., obese patients or binge eaters). The study involving obese patients98 used the stop-signal task. While similar SSRTs were found, obese patients showed less brain activation than controls in major parts of the inhibitory control network (i.e., right SFG, left IFG, bilateral MFG, insula, IPL, cuneus, right occipital regions and left MTG). In the study by Lock and colleagues,99 similar accuracy levels were found during a go/no-go task, while participants with binge eating behaviour had more brain activation associated with inhibitory control than controls in brain regions critically involved in inhibitory control, such as the right DLPFC, right ACC, bilateral precentral gyri, bilateral hypothalamus and right MTG.
Summary
Event-related potential findings in excessive Internet users showed reduced N2 amplitudes in 2 studies, suggesting a deficit in the conflict detection stage of the inhibition process. In contrast, N2 amplitudes in people with excessive gaming behaviour were enhanced in a parietal cluster. One fMRI study in individuals with pathological gambling showed hypoactivation associated with inhibitory control in the dACC, while a second fMRI study showed that inhibitory control and related brain activation may be boosted by a gambling-related context. Findings of the 2 fMRI studies in people with excessive eating behaviour seem to partly contradict each other. While neither study showed behavioural deficits in inhibitory control, 1 study showed hyperactivation in patients whereas the other showed hypoactivation in substantial parts of the inhibitory control network. Clearly, more neuroimaging studies in populations with excessive addiction-like behaviours are necessary.
Error processing
Error processing in individuals with nicotine dependence
Two ERP and 2 fMRI studies have examined error processing in individuals with nicotine dependence. Franken and colleagues100 found that Flanker task performance and ERN amplitudes for incorrect trials were not impaired in individuals with nicotine dependence after 1 hour of smoking abstinence. However, Pe amplitudes were lower in these individuals than in controls. These findings may indicate that initial error detection in individuals with nicotine dependence is intact but that more conscious evaluation of errors may be less distinct in this group. Luijten and colleagues101 used a similar task in a study of individuals with nicotine dependence after 1 hour of abstinence, but also included smoking cues. Both ERN and Pe amplitudes were lower in those with nicotine dependence than controls. In addition, smokers displayed less post-error slowing than controls. Results of this study and that of Franken and colleagues100 suggest that initial error detection may be specifically compromised in individuals with nicotine dependence when limited cognitive resources are available for error monitoring (e.g., during exposure to smoking cues). On the other hand, the more conscious processing of errors may be generally less distinct in individuals with nicotine dependence.
An fMRI study in which participants performed a stop-signal task showed less error-related activation in individuals with nicotine dependence than controls in the dACC coupled with increased activation in an anterior region of the dorsomedial prefrontal cortex (DMPFC).62 Using a go/no-go task, Nestor and colleagues63 found that nonabstaining individuals with nicotine dependence, as compared with controls, made more errors accompanied by reduced brain activation after performance errors in the right SFG and left STG, whereas no difference was found in either the ACC or insula. This study also included a group of ex-smokers who were abstinent for at least 1 year and showed enhanced error-related activity in the ACC, left insula, bilateral SFG, right MFG, left cerebellum, left MTG, bilateral STG and bilateral parahippocampal gyrus (PHG) relative to individuals with nicotine dependence and controls. These findings suggest that more elaborate neural monitoring of errors may increase the probability to quit smoking or that the deficits in individuals with nicotine dependence are reversible.
Summary
Results from the 2 ERP studies suggest that initial error detection may be less effective in individuals with nicotine dependence during more cognitively challenging situations, whereas the more conscious evaluation of errors may also be compromised in affectively neutral conditions. Hypoactivation in the ACC in reaction to errors was found in 1 of the 2 fMRI studies in individuals with nicotine dependence. Further research should clarify under which conditions neural deficits associated with error processing are present in these individuals.
Error processing in individuals with alcohol dependence
Two ERP studies and 1 fMRI study have investigated error processing in abstinent patients with alcohol dependence. Padilla and colleagues102 and Schellekens and colleagues103 investigated ERN (but not Pe) amplitudes in abstaining individuals with alcohol dependence evoked by errors on a Flanker task. The alcohol dependence group in the study by Padilla and colleagues102 performed the task as accurately as the control group but showed increased ERN amplitudes, suggesting enhanced monitoring of performance errors. However, this may not be specific for errors in this study, since the alcohol dependence group also showed increased amplitudes for correct trials. Another ERP study in individuals with alcohol dependence found increased ERN amplitudes specifically for errors in patients with alcohol dependence relative to controls.103 In addition, these alcohol-dependent patients showed increased error rates for congruent trials. Interestingly, when individuals with alcohol dependence and comorbid anxiety disorders were compared with those without anxiety disorders, ERN amplitudes were larger in the anxiety subgroup. Enhanced ERN amplitudes in highly anxious individuals is in line with theories suggesting that internalizing psychopathology is associated with increased monitoring of performance errors.104 In line with ERP findings, an fMRI study by Li and colleagues75 showed increased error-related brain activation in individuals with alcohol dependence relative to controls in a stop-signal task in the right ACC, bilateral MFG and bilateral SFG as well as in regions outside the error processing network (i.e., the bilateral MTG, SPL, right central culcus and right superior and middle occipital gyrus).
Summary
It appears that the processing of errors is enhanced in abstaining individuals with alcohol dependence, as ERN amplitudes and error-related ACC activation were increased. Currently, none of the ERP studies in individuals with alcohol dependence evaluated Pe amplitudes; therefore, no information is available regarding more conscious processing of errors in this group.
Error processing in individuals with cannabis dependence
No ERP studies and only 1 fMRI study investigating error processing in individuals with cannabis dependence were identified.81 In the fMRI study, participants were asked to press a button in a go/no-go task when they noticed that they made a mistake, such that aware and unaware errors could be evaluated separately. For aware errors, activation in regions critical for error processing was similar in non–treatment seeking individuals with cannabis dependence and controls, whereas cannabis-dependent individuals showed more error-related brain activation in the bilateral precuneus and left putamen, caudate and hippocampus. The proportion of errors in cannabis-dependent individuals and controls was similar; however, the cannabis-dependent individuals were less often aware of their errors. In addition, cannabis-dependent individuals, but not controls, showed less activation in the right ACC, bilateral MFG, right putamen and IPL for unaware errors than aware errors. The difference in error-related ACC activity for aware and unaware errors was positively associated with reduced error awareness.
Summary
More fMRI studies are needed to confirm less pronounced error awareness in cannabis users. Also, ERP studies should evaluate whether the initial automatic stage of error processing may also be compromised and should replicate less distinct error awareness in individuals with cannabis dependence by evaluating Pe amplitudes.
Error processing in individuals with stimulant dependence
Three ERP studies investigated error processing in individuals with cocaine dependence.7,85,105 No studies in populations using other stimulants were identified. Participants in the study by Franken and colleagues7 performed a Flanker task. Event-related potential findings showed that both the initial automatic processing of errors and the later more conscious processing of errors is less pronounced in abstaining individuals with cocaine dependence than controls, as both ERN and Pe amplitudes were attenuated. Furthermore, participants with cocaine dependence committed more errors than controls. More specifically, they committed more errors after an error on the previous trial, which suggests that behavioural adaptation was suboptimal. Sokhadze and colleagues85 and Marhe and colleagues105 confirmed increased error rates and reduced ERN amplitudes in individuals with cocaine dependence relative to controls performing, respectively, a combined Flanker and go/no-go task in active users and a classic Flanker task in cocaine-dependent patients in the first few days of detoxification. Neither study investigated Pe amplitudes. Importantly, reduced ERN amplitudes were also shown to be predictive for increased cocaine use at 3-month follow-up.105
Two fMRI studies in individuals with cocaine dependence investigated brain activation associated with error processing employing a go/no-go87 and a stop-signal task.89 Error-related hypoactivation was found in those who were actively using cocaine compared with controls in the ACC, right MFG, left insula and left IFG. In addition, individuals with cocaine dependence committed more errors during task performance. In line with ERP findings, Luo and colleagues89 showed that reduced error-related dACC activation in abstaining individuals with cocaine dependence was associated with relapse rates 3 months later in both men and women, while sex-specific effects were found in the thalamus and the left insula.
Summary
Both ERP and fMRI studies show less error-related brain activation in individuals with cocaine dependence than controls, especially in regions critical for optimal error processing, such as the ACC, insula and IFG. Lower ERN and Pe amplitudes in individuals with cocaine dependence compared to controls suggest that problems with error processing may emerge as both a consequence of deficits in initial error detection as well as from deficits in the more conscious evaluation of performance errors. Reduced ERN amplitudes and error-related dACC activation were associated with relapse at 3-month follow-up.
Error processing in individuals with opiate dependence
We identified no ERP studies and only 1 fMRI study that investigated error processing in abstaining individuals with opiate dependence.106 It was found that individuals with opiate dependence made more errors in a go/no-go task and that error-related activation in the ACC was reduced compared with activation in controls. Furthermore, an association between ACC activation and behavioural performance in individuals with opiate dependence was lacking, whereas this brain–behaviour correlation was present in controls.
Summary
Neural deficits in error-related brain activation in the ACC in individuals with opiate dependence were found in an fMRI study. Obviously, more fMRI and ERP studies are needed to confirm differences in these patients.
Error processing in individuals with behavioural addictions
We identified only 1 ERP study in the domain of behavioural addictions that showed increased errors rates for no-go trials in people with excessive gaming behaviour compared with controls.96 Lower ERN amplitudes and no differences in Pe amplitudes were found in participants with excessive gaming for error trials, suggesting that initial error processing in excessive gamers may be less pronounced than in controls, whereas error awareness may not be related to increased error rates. The only fMRI study that investigated error processing in the context of behavioural addictions showed that error-related brain activation in the dACC on the stop-signal task was lower in individuals with pathological gambling behaviour than controls, whereas task performance was intact.62 This finding suggests a less pronounced monitoring of errors in the pathological gambling group in the most important region for error processing.
Summary
Both studies investigating error processing showed less processing of errors in individuals with excessive addiction-like behaviours, thereby resembling findings in individuals with substance dependence. Additional fMRI and ERP studies are needed to replicate these findings and extend them to other groups showing behavioural addictions.
Discussion
Summary of results
The present review provides an overview of ERP and fMRI studies that have addressed inhibitory control and error processing in individuals with substance dependence and in individuals showing proposed behavioural addictions. The ERP studies of inhibitory control, as operationalized using go/nogo and stop-signal paradigms, have found deficits in N2 and P3 amplitudes in individuals with addictions. Of the studies that evaluated N2 amplitudes (n = 7), most (n = 5) showed lower N2 amplitudes in individuals with addictions than controls (for an example, see the Appendix, Fig. S1, at jpn.ca), suggesting that deficits in inhibitory control in individuals with addictions could be caused by problems with early cognitive processes, such as conflict detection. Results of studies on P3 amplitudes (n = 11) are inconsistent. Some studies showed no differences between individuals with addictions and controls (n = 5), whereas other studies showed lower (n = 5) or higher (n = 1) P3 amplitudes in those with addictions. Therefore no clear conclusions can be formulated with regard to the P3. Complementary to findings of less pronounced N2 amplitudes, several fMRI studies (n = 13 of 16) found hypoactivation associated with inhibitory control in individuals with addictions, mainly in the ACC, IFG and DLPFC, but also in the inferior and superior parietal gyri (Fig. 1). From these findings, it can be concluded that substantial parts of the network underlying inhibitory control are dysfunctional in individuals with addictions. Of note, differences in brain activation associated with inhibitory control were also found outside the inhibitory control neural network, signifying that individuals with addictions may use different strategies to implement inhibitory control.
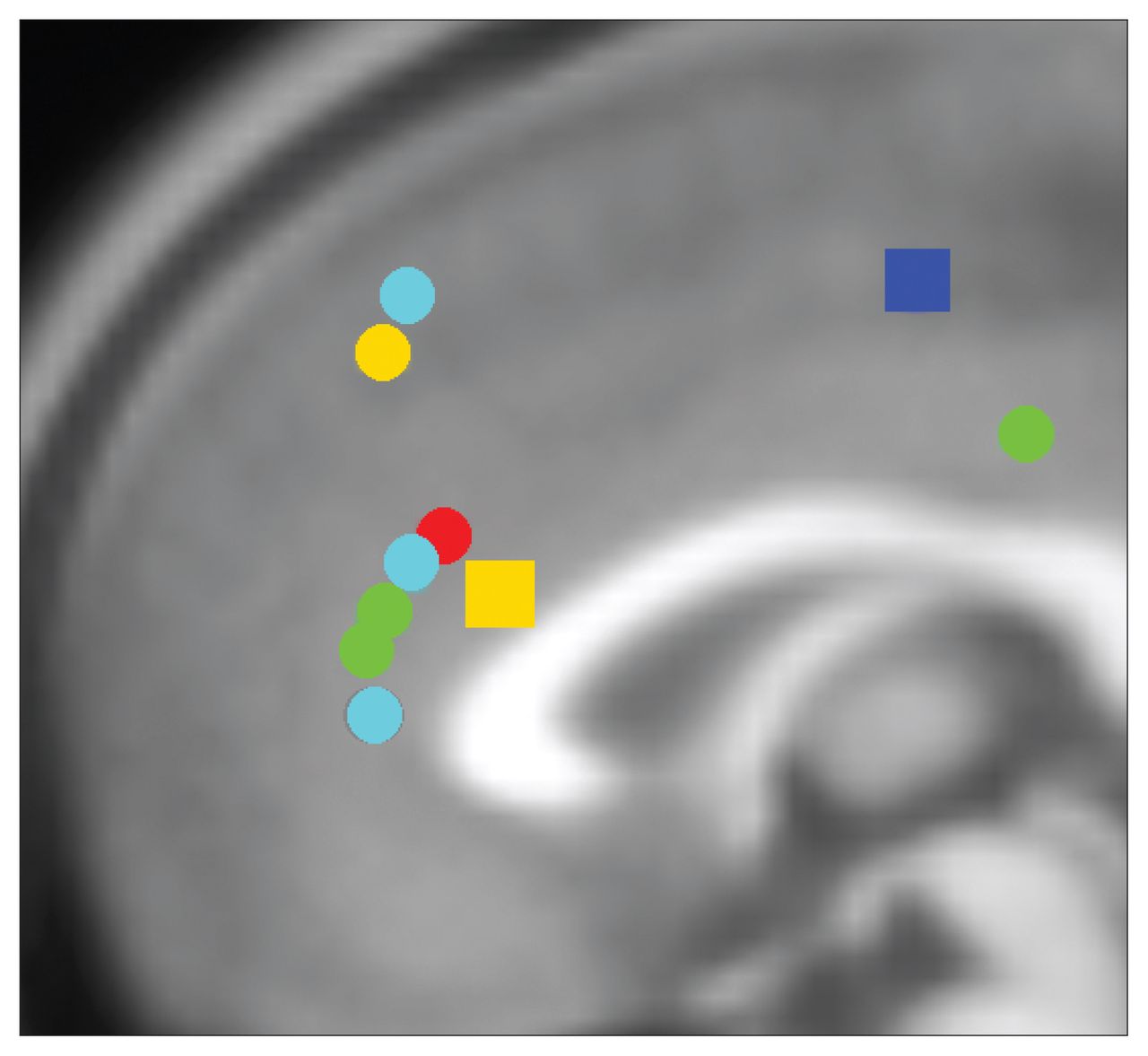
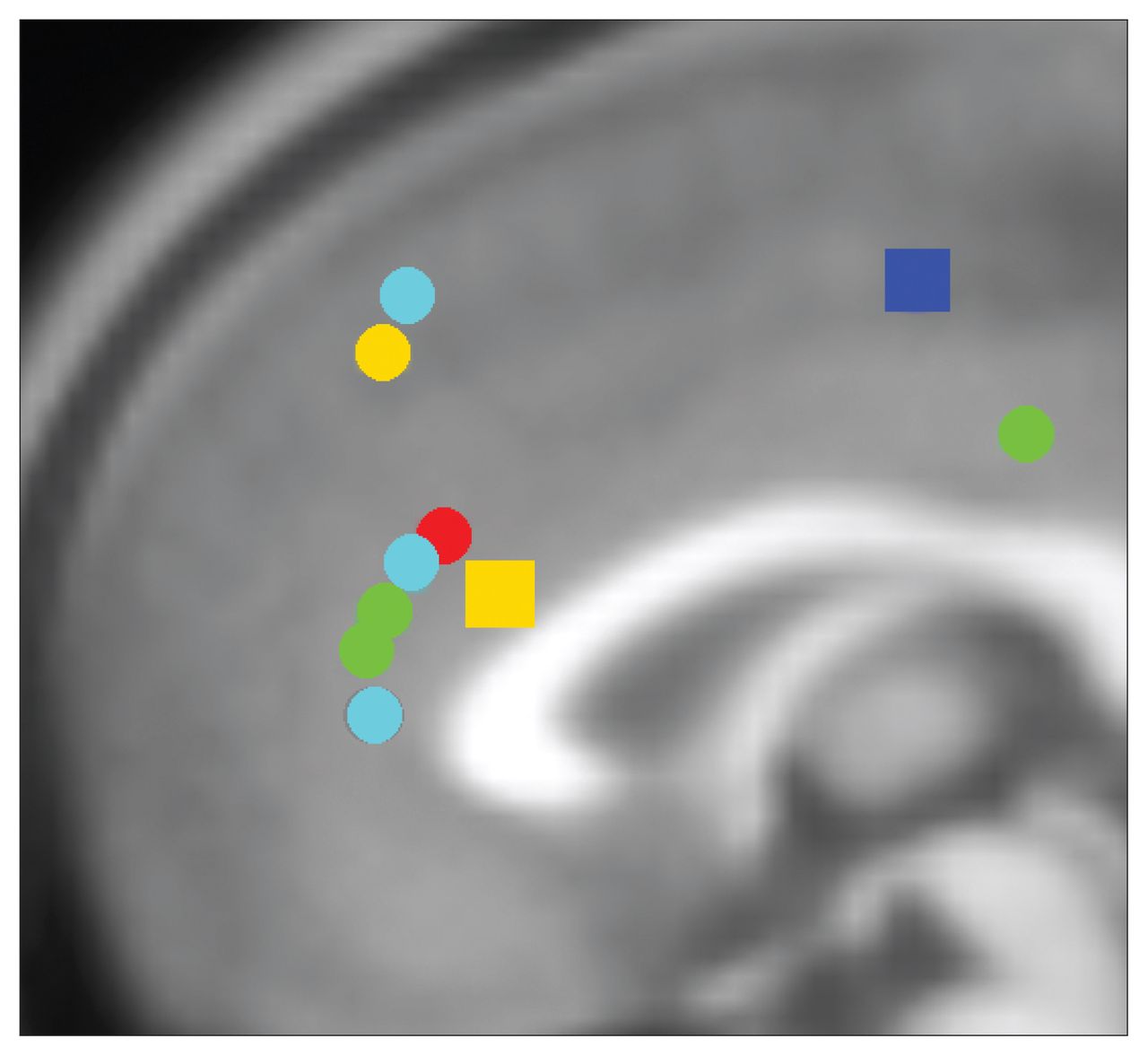
Summary of anterior cingulate dysfunction in individuals with addictions for inhibitory control. Circles represent hypoactivation and squares hyperactivation for inhibitory control in individuals with addictions relative to controls. Of note, 6 studies included in this review reporting group differences are not displayed in this figure, as results did not indicate group differences in the anterior cingulate cortex for inhibitory control–related brain activation. Red: opioid dependence; green: cocaine dependence; dark blue: cannabis dependence; light blue: nicotine dependence; yellow: behavioural addictions. Locations are based on reported Talairach or Montreal Neurological Institute (MNI) coordinates in studies reporting group differences included in this review. Talairach coordinates were converted to MNI using the GingerALE toolbox. Foci of activation were projected onto the midline for ease of viewing.
Error-related hypoactivation in individuals with addictions in the ACC, the most critical area for error processing, was found in most (n = 6 of 7) fMRI studies (Fig. 2), while hypoactivation associated with error processing was also reported in other regions, such as the superior and inferior frontal gyri and insula. The ERP findings confirm and complement fMRI findings. Lower ERN amplitudes in individuals with addictions relative to controls were observed (n = 5 of 8), thereby confirming initial error-detection deficits in individuals with addictions (see the Appendix, Fig. S2, for an example of ERN and Pe findings). Given that the ACC is the neural generator of the ERN,8,48,49 both ERN and fMRI findings suggest that ACC dysfunction could be a biomarker for error processing deficits in individuals with addictions. Importantly, lower ERN amplitudes and hypoactivation in the ACC was associated with relapse in 2 longitudinal studies.89,105 Pe findings complement fMRI findings by providing information on the time frame of error processing deficits. Lower Pe amplitudes in individuals with substance dependence compared with controls were observed (n = 3 of 4) and suggest that, besides the initial error detection, more conscious processing of errors may also be compromised. This is a particularly interesting finding as it may be associated with impaired insight in behaviour, a topic that recently attracted more attention in the field of addiction.107

Summary of anterior cingulate dysfunction in individuals with addictions for error processing. Circles represent hypoactivation and squares hyperactivation for error processing in individuals with addictions relative to controls. Of note, 1 study included in this review that reports group differences is not displayed in this figure as results did not indicate group differences in the anterior cingulate cortex for error-related brain activation. Red: opioid dependence; green: cocaine dependence; purple: alcohol dependence; light blue: nicotine dependence; yellow: pathological gambling. Locations are based on reported Talairach or Montreal Neurological Institute (MNI) coordinates in studies reporting group differences included in this review. Talairach coordinates were converted to MNI using the GingerALE toolbox. Foci of activation were projected onto the midline for ease of viewing.
Two findings in the present review constitute an exception to the conclusions discussed. First, fMRI findings in cannabis users show hyper- instead of hypoactivation with regard to inhibitory control in brain regions critically involved in inhibitory control, including the pre-SMA, DLPFC, insula and IPG. Hyperactivation associated with inhibitory control in cannabis users can be interpreted as increased neural effort to reach control sample levels of behaviour performance (i.e., no behavioural deficits were found in these individuals). Another explanation for hyperactivation in this population is the comparatively young age of cannabis users in both fMRI studies relative to other studies in individuals with substance dependence.81,82 In addition, participants in the study by Tapert and colleagues82 abstained from cannabis use for 28 days, which is longer than in most other studies, suggesting that brain activation may change as a function of abstinence duration.108
The ERP and fMRI findings regarding error processing in individuals with alcohol dependence constitute the second exception on the generally observed error-related hypoactivation in individuals with addictions. In contrast to other populations with addictions, those with alcohol dependence show enhanced error processing, as reflected by enlarged ERN amplitudes and increased error-related activation in the ACC.75,102,103 Findings in the study by Schellekens and colleagues103 provide a possible explanation for enhanced error processing in alcohol-dependent individuals, as ERN amplitudes were larger in highly anxious individuals than in less anxious individuals. This suggests that the often observed co-morbid internalizing psychopathology (i.e., anxiety-related disorders) in individuals with alcohol dependence109,110 may be responsible for the enhanced error processing. An overview of ERN findings confirms that internalizing psycho-pathology is associated with larger ERN amplitudes, whereas externalizing psychopathology is associated with less pronounced ERN amplitudes.104
A secondary goal of our review was to evaluate differences and similarities in inhibitory control and error processing between substance dependence and other addictive behaviours. Similar findings to those observed in individuals with substance dependence were found in people with pathological gambling and excessive eating, gaming and Internet use. For example, hypoactivation in the ACC for both inhibitory control and error processing was found in individuals with pathological gambling behaviour,62 which resembles the most often observed finding in individuals with substance dependence. However, contradictory findings have also been identified in those with excessive gaming behaviour (e.g., enlarged N2 amplitudes) and excessive eating behaviour (i.e., 1 fMRI study in the domain of inhibitory control showed hypoactivation during an inhibition task, whereas the other showed hyperactivation). In conclusion, some similarities between individuals with substance dependence and those showing addictive behaviours were identified; however, there remains insufficient neuroimaging research in these populations, and current findings are inconclusive.
Integrative model
The integration of ERP and fMRI findings for both inhibitory control and error processing results in the observation that most consistent findings in individuals with addictions are all related to dysfunction of the dACC. Both the N2 and the ERN have their neural origin in the dACC,111 and dACC dysfunction was the most consistent fMRI finding for both inhibitory control and error processing. This suggests that a shared dysfunction of the dACC may contribute to deficits in both inhibitory control and error processing. An influential theory concerning the function of the dACC suggests that conflict monitoring is the core function of the dACC,8,112 thereby explaining its crucial role in many different cognitive functions. This theory is supported by the finding that conflict-related activation in the dACC precedes increased activation in the DLPFC on the next trial, showing that the dACC precedes adjustment in activation in other brain regions that implement cognitive control.59 This conflict monitoring function of the dACC may be a critical function for both inhibitory control and error processing. For inhibitory control, a conflict between the automatic response tendency and the long-term goal needs to be detected to inhibit behaviour. Error processing and conflict monitoring may even be more tightly related, probably in a reciprocal way. To be able to process errors during ongoing behaviour, the monitoring of conflict is crucial to signal the difference between the actual response and the representation of the correct response. On the other hand, an optimal processing of performance errors is necessary for learning and conflict monitoring in future behaviour, thereby illustrating a possible reciprocal relationship between conflict monitoring and error processing. This reciprocal association with conflict monitoring (Fig. 3) suggests that error processing deficits may indirectly influence other functional domains of cognitive control, including inhibitory control.113 Altogether, we propose that disturbed conflict monitoring in the dACC represents a core deficit in individuals with addictions underlying the observed deficits in error processing and inhibitory control (Fig. 3). Notably, this idea of conflict monitoring as a common deficit in dACC functioning in individuals with addictions may generalize to other domains of cognitive control, including feedback processing, attentional monitoring and salience detection. In line with this idea, some of these functions, such as salience detection measured in oddball paradigms, have previously been shown to be impaired in individuals with addictions,114 whereas other functions, such as attentional monitoring, constitute a crucial part of many cognitive control functions, including inhibitory control. Given the proposed role of the IFC in go/no-go and stop-signal task paradigms, the observed IFG deficits in individuals with addictions during these tasks may reflect reduced attentional monitoring capacities.19,20,115 Based on the postulated model it can be expected that improvement of dACC functioning, either through direct neuromodulation or indirect behavioural therapies would result in increased control over addictive behaviours. Another hypothesis based on the current model would be that interventions targeting conflict monitoring or error processing would concomitantly result in improvements in inhibitory control, while this would not necessarily work in the opposite direction.

Summary and integrative model of neural deficits in error processing and inhibitory control in individuals with addictive behaviours. The event-related potential components and brain regions listed in the boxes are those showing the most consistent neural deficits in individuals with addictions in error processing and inhibitory control. As the most consistent findings are related to dorsal anterior cingulate cortex (dACC) dysfunction, we suggest that problems with conflict monitoring, a critical function of the dACC, involved in both inhibitory control and error processing, may trigger the loss of control in individuals with addictions. The grey arrow indicates that the function of this highly interrelated system is decreased in individuals with addictions compared with healthy controls. It further illustrates the negative impact of this dysfunctional system over addiction-related behaviours. The arrows in the model symbolize the relationships between error-processing and inhibitory control with conflict monitoring. The dotted arrow represents the modulatory effect of error-related dACC activation on connected brain regions. DLPFC = dorsolateral prefrontal cortex; ERN = error-related negativity; IFG = inferior frontal gyrus; PG = parietal gyrus.
Limitations
It is crucial to note that inconsistencies in findings within and among included studies were evident. For example, brain and behavioural findings were not always consistent and individuals with addictions showed hyper- instead of hypo-activation associated with inhibitory control or error processing in some studies. Generally, the interpretation of hypo-versus hyperactivation in ERP and fMRI studies in clinical populations relative to controls remains equivocal. Behavioural findings, such as less accurate task performance or reaction time differences, are key to guide the interpretation of hypo- or hyperactivation. Although speculative, a possible explanation for hypoactivation without behavioural deficits is that brain activation may be a more sensitive measure to detect abnormalities in individuals with addictions.5,116 In this context it would be interesting to investigate associations between the amount of substance use or the level of dependency and the extent of hypoactivation. On the other hand, hyperactivation coupled with intact behavioural performance is often interpreted as increased neural effort or the use of alternative cognitive strategies to achieve normal levels of behavioural performance.117
Inconsistencies in results are probably owing to differences in methodology, such as patient selection, specifications of task paradigms, data acquisition and analysis techniques. Although we report several patient characteristics in Table 1, it is a limitation of the present review that the effects of these characteristics on neuroimaging results could not be evaluated owing to the large variability and the limited number of studies. In particular, abstinence duration has been shown to alter cognitive control and associated brain function.118 Therefore, longitudinal studies are clearly needed to unravel the developmental trajectory of cognitive deficits after prolonged periods of drug abstinence. A further limitation is that it was unclear in some studies whether the researchers adequately controlled for nicotine use. As the current review clearly showed differences in inhibitory control and error processing and related brain activation in smokers versus nonsmokers, nicotine use should be taken into account in studies of other populations with addictions.
Another limitation of the present review is the small number of studies included for some substances of abuse, which hampered firm conclusions in those groups. More studies are needed, particularly in individuals with opiate and cannabis dependence and in individuals showing excessive addiction-like behaviours. In addition, we recommend that both ERN and Pe, or N2 and P3 amplitudes be evaluated in a single study to provide optimal information regarding the time frame of cognitive control deficits.
With regard to task paradigms, it is a strength of the present review that we selected only those task paradigms that most closely reflect inhibitory control and error processing (i.e., go/no-go, stop-signal and Flanker task), thereby reducing variability in results owing to the different cognitive processes needed for task performance. On the other hand, the narrow focus may be considered a limitation, as results cannot be generalized to other cognitive domains or task paradigms. Studies using the Stroop task, for example, were excluded because the Stroop task is known to evoke cognitive processes, such as conflict resolution, response selection and attention23,24 as well as different ERP components compared with go/no-go and stop-signal paradigms.119–121 Nevertheless, some findings in fMRI and positron emission tomography studies using the classic colour–word Stroop task are in line with the present findings.122–124 Even with the strict selection of task paradigms, there is still variance in outcomes within go/no-go and stop-signal paradigms, which contributes to inconsistencies in results across studies. Differences in analysis techniques may further induce inconsistencies in results. For fMRI studies, whole brain versus regions of interest analyses and different methods to correct for multiple comparisons are major sources of variance, and so is the use of different within-subject contrasts for subsequent between-subject analyses (e.g., stop correct minus go v. stop correct minus stop error). Task design and analysis techniques should become much more standardized to reduce inconsistencies in results. This is also a prerequisite if these paradigms will eventually be implemented in clinical practice.
Treatment implications and future research directions
Contemporary effective treatments for addiction involve pharmacotherapy, cognitive behaviour therapy and contingency management.125–127 Nevertheless, relapse rates are still high, so there is ample room for improvement. Several treatment targets based on the findings in this review merit further investigations. First, it has been shown that inhibitory control capacities and underlying neural networks could be trained to increase behavioural control.128 A second possibility to increase inhibitory control is the direct training of hypoactive brain regions, such as the ACC, IFG and DLPFC, using neuromodulation techniques.129–131 Specific medications with the aim to enhance cognitive functions may be another treatment intervention to increase cognitive functioning.132 More research into these clinical applications is needed to explore which of these potential treatment strategies may eventually be effective in the reduction of addictive behaviours.
Cognitive control capacities can also be used in clinical practice to guide treatment strategies according to individual needs. It has been shown that deficits in cognitive control are associated with a reduced capacity to recognize problems with substance abuse, lower motivation to enter treatment and treatment dropout.133,134 Berkman and colleagues66 showed that individual differences in activation in the inhibitory control network are linked to the ability to inhibit craving in daily life to prevent smoking. These and other recent findings135 highlight the need to monitor cognitive control capacities during treatment and may be used to identify individuals with addictions who are more vulnerable to relapse.
One of the most important remaining questions is that of causality. It is not yet known whether neural deficits associated with inhibitory control and error processing in individuals with addictions predispose them to substance use or whether they are a consequence of substance use. Interestingly, a recent study provided evidence for the ERN as a possible endopheno-type for addiction,136 as ERN amplitudes were lower in high-risk offspring than adolescents with a normal risk.
Conclusion
This review systematically evaluated ERP and fMRI findings concerning inhibitory control and error processing in individuals with substance dependence and individuals showing excessive addiction-like behaviours. The combined evaluation of ERP and fMRI offers new insights and future research directions. Overall, results show that addiction is associated with neural deficits related to inhibitory control and error processing. The most consistent findings were lower N2, ERN and Pe amplitudes and hypoactivation in the dACC, IFG and DLPFC in individuals with addictions compared with controls. We propose an integrative model suggesting that dACC dysfunction in conflict monitoring may be a core neural deficit underlying addictive behaviours. Finally, similarities between individuals with substance dependence and individuals showing addiction-like behaviours were identified, but evidence for neural deficits in the domains of inhibitory control and error processing in the latter population is scarce and inconclusive.
Acknowledgements
This study was supported by a grant of the Netherlands Organization for Scientific Research (NWO; VIDI grant number 016.08.322). The funding organization had no role in preparation of the manuscript or decision for publication. The authors have no competing interests to declare.
Footnotes
Competing interests: None declared.
Contributors: All authors designed the study, acquired and analyzed the data and approved the final version to be published. M. Luijten and M.W.J. Machielsen wrote the article, which D.J. Veltman, R. Hester, L. de Haan and I.H.A. Franken reviewed.
- Received March 25, 2013.
- Revision received June 25, 2013.
- Accepted July 10, 2013.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
Article tools



Related Articles
Cited By...
- Caffeinated soda intake in children is associated with neurobehavioral risk factors for substance misuse
- Recovery of inhibitory control prefrontal cortex function in inpatients with heroin use disorder: a 15-week longitudinal fMRI study
- The Neural Signature of Impaired Inhibitory Control in Individuals with Heroin Use Disorder
- The neural signature of impaired inhibitory control in individuals with heroin use disorder
- An Examination of the Relationship Between Perfectionism and Neurological Functioning
- Electrophysiological correlates of proactive and reactive inhibition in a modified visual Go/NoGo task
- Event-related and oscillatory signatures of response inhibition: A magnetoencephalography study with subclinical high and low impulsivity adults
- An Integrated Multimodal Model of Alcohol Use Disorder Generated by Data-Driven Causal Discovery Analysis
- Brain networks underlying vulnerability and resilience to drug addiction
- Neural correlates of response inhibition and performance monitoring in binge watching
- Nicotine dependence (trait) and acute nicotinic stimulation (state) modulate attention but not cognitive control: converging fMRI evidence from Go-Nogo and Flanker tasks
- Brain responses to anticipating and receiving beer: Comparing light, at-risk, and dependent alcohol users