

Abstract
Background: Social anxiety disorder is characterized by intense fear and avoidance of social interactions and scrutiny by others. Although alterations in attentional control seem to play a central role in the psychopathology of social anxiety disorder, the neural underpinnings in prefrontal brain regions have not yet been fully clarified.
Methods: The present study used functional MRI in participants (age 18–50 yr) with social anxiety disorder (n = 42, 31 female) and without (n = 58, 33 female). It investigated the interrelation of the effects of social anxiety disorder and early-life adversity (a main environmental risk factor of social anxiety disorder) on brain activity during an attentional control task. We applied DNA methylation analysis to determine whether epigenetic modulation in the gene encoding the glucocorticoid receptor, NR3C1, might play a mediating role in this process.
Results: We identified 2 brain regions in the left and medial prefrontal cortex that exhibited an interaction effect of social anxiety disorder and early-life adversity. In participants with low levels of early-life adversity, neural activity in response to disorder-related stimuli was increased in association with social anxiety disorder. In participants with high levels of early-life adversity, neural activity was increased only in participants without social anxiety disorder. NR3C1 DNA methylation partly mediated the effect of social anxiety disorder on brain activity as a function of early-life adversity.
Limitations: The absence of behavioural correlates associated with social anxiety disorder limited functional interpretation of the results.
Conclusion: These findings demonstrate that the neurobiological processes that underlie social anxiety disorder might be fundamentally different depending on experiences of early-life adversity. Long-lasting effects of early-life adversity might be encoded in NR3C1 DNA methylation and entail alterations in social anxiety disorder–related activity patterns in the neural network of attentional control.
Introduction
Social anxiety disorder is a psychiatric disorder characterized by excessive anxiety in social situations, centring around the fear of embarrassment and negative evaluation. A key element in the psychopathology of social anxiety disorder is a distortion in information processing that involves alterations of attentional control with an unintentional deployment of attentional resources toward disorder-related stimuli.1 In recent years, many studies have investigated the neural underpinnings of anxiety-related stimulus processing (e.g., emotional faces,2 self-referential comments3 or social anxiety disorder–related words4,5) in social anxiety disorder. Perception of such stimuli consistently involves hyperactivity in the amygdala and insular cortex,6 but also altered activation of the anterior cingulate, the parietal cortex, the hippocampus and the fusiform gyrus.7 Alterations in activity in the ventrolateral cortex, the dorsolateral prefrontal cortex and the mediofrontal cortex have often been reported in association with reduced ability to engage in top–down attentional control in social anxiety. Unaccountably, however, findings in the frontal cortex were inconsistent because both decreased and increased hemodynamic activation were observed.7,8
A meta-analysis identified the inferior frontal junction in the lateral prefrontal cortex as one of the key regions of the attentional system.9 It also reported that the anterior insula, the anterior midcingulate cortex and presupplementary motor area were consistently associated with attentional control. Therefore, certain convergent brain regions have been found to be dysregulated in social anxiety disorder that generally play a key role in attentional control.
One of the most prevalent contextual factors of social anxiety disorder is early-life adversity.10 The early life is thought to be a particularly sensitive period, because the brain and its neural circuits are still developing. Disruptions by adverse events during this time window can have long-lasting behavioural and neurobiological effects. Behaviourally, alterations in attentional control11 and emotion processing11,12 have been reported; neuronally, early-life adversity has most consistently been associated with increased activity in the amygdala in response to threat-related stimuli, increased activation of the anterior cingulate cortex during emotion regulation, and altered lateral prefrontal activity in relation to executive functions. However, for the latter, the direction of effect varies among studies.13,14
Based on this strong congruence of brain regions that are dysregulated in association with both social anxiety disorder and early-life adversity — and on accumulating evidence that early-life adversity may represent an unrecognized confounding moderator in many psychiatric neuroimaging studies14 — the goal of the present study was to investigate the interrelation of the effects of social anxiety disorder and early-life adversity on brain activity patterns.
The early life not only represents a period of increased neuroplasticity but also appears to be characterized by substantial developmental epigenetic plasticity.15 Epigenetic modifications present a mechanism for how environmental factors such as early-life adversity might affect gene regulation — for example, through DNA methylation.16 Primarily based on candidate gene studies motivated by work in rodents, the DNA methylation pattern of the NR3C1 gene has consistently been associated with both early-life adversity and anxiety disorders, including social anxiety disorder.17,18 NR3C1 encodes the glucocorticoid receptor, which plays a pivotal role in the body’s stress response system along the hypothalamic–pituitary–adrenal axis.19 Although increased NR3C1 DNA methylation in association with early-life adversity has been observed in most studies, some have reported no associations or an opposite direction of effect.17
We investigated the interactions of social anxiety disorder and early-life adversity at the level of hemodynamic brain activation, focusing on attentional control. In particular, we examined whether the experience of early-life adversity might moderate activity differences associated with social anxiety disorder and, hence, might account for previously observed inconsistencies in brain activation patterns related to social anxiety disorder. We applied DNA methylation analysis to determine whether the DNA methylation of the NR3C1 gene might serve as a potential mediator of the effects of early-life adversity on brain function related to social anxiety disorder.
Methods
Participants
We screened 119 native German-speaking participants (age 18–50 yr) of European descent for study inclusion. All participants underwent the German version of the Structured Clinical Interview for DSM-IV.20 Of those, 48 participants had a primary diagnosis of social anxiety disorder. We evaluated the severity of participants’ social anxiety using the Liebowitz Social Anxiety Scale.21 Six individuals with a diagnosis of social anxiety disorder but a low Liebowitz Social Anxiety Scale score (< 3522) and 13 without a diagnosis of social anxiety disorder but a high Liebowitz Social Anxiety Scale score (≥ 35) were not included in our sample because of this inconsistency. We assessed early-life adversity using the Childhood Trauma Questionnaire, which measures 5 types of maltreatment occurring up to adolescence: emotional and physical neglect, and emotional, physical and sexual abuse. Participants who had at least a moderate score in 1 of the 5 categories (i.e., ≥ 15 for emotional neglect, ≥ 13 for emotional abuse, ≥ 10 for physical abuse or neglect, ≥ 8 for sexual abuse) were classified as having a high level of early-life adversity.23,24 Thus, 4 groups emerged: controls with low (n = 34) and high (n = 24) levels of early-life adversity, and participants with social anxiety disorder and low (n = 21) and high (n = 21) levels of early-life adversity. We assessed general anxiety using the State–Trait Anxiety Inventory25 and depressive symptoms using the Beck Depression Inventory.26 A sample description is detailed in Appendix 1, Table S1, available at jpn.ca.
All participants provided written informed consent before study inclusion. The study was performed in accordance with the Declaration of Helsinki in its latest version and approved by the University of Tübingen local ethics committee.
Stimuli and task
Participants conducted an emotional counting Stroop task. Stimuli comprised 60 words from 4 different categories (15 each): neutral, social anxiety disorder–related, fear-related and generally negative words. Each stimulus was presented in 3 conditions (i.e., repeated 2, 3 or 4 times), resulting in 180 stimulus events. Each stimulus was presented for 500 ms. The mean intertrial interval was 6 s (range 4.5–7.5 s). For further details, see Appendix 1.
Image acquisition
We acquired MRI data using a 3 T scanner (PRISMA, Siemens). We recorded structural magnetization-prepared rapid gradient echo T1-weighted images using the following parameters: 208 slices, repetition time 2400 ms, echo time 2.22 ms, voxel size 0.8 × 0.8 × 0.8 mm3. For functional imaging, we used multiband echo-planar imaging sequences with the following parameters: 72 slices, 2 mm thickness, repetition time 1500 ms, echo time 34 ms, voxel size 2 × 2 × 2 mm3, 850 volumes. For further details, see Appendix 1.
Experimental procedure
Study participation included 2 appointments within 1 week. In the first session, participants were screened for current or previous psychiatric disorders using the Structured Clinical Interview for DSM-IV;20 completed several questionnaires related to basic demographic data, anxiety, depression and early-life adversity; and provided a blood sample. In the second session, eligible participants took part in the functional MRI experiment.
Behavioural and sample analysis
We analyzed behavioural data using SPSS Statistics (version 26.0; IBM), and we created graphics using R (version 3.5.1).27 We analyzed the effects of social anxiety disorder, early-life adversity and their interaction on the characteristics of the sample using a univariate analysis of variance (ANOVA) with the dichotomous factors social anxiety disorder and early-life adversity, as well as their interaction. Because of technical problems, responses were not recorded for 6 participants. We excluded reaction times for incorrect trials, trials with multiple responses and trials with a reaction time more than 2 standard deviations from the mean reaction time that was within each word category (exclusion of about 6% of trials). We analyzed the mean reaction time for neutral and social anxiety disorder–related words in an ANOVA, with word category as the repeated-measures factor; early-life adversity and social anxiety disorder as between-subject factors; and age and sex as covariates. Similarly, we analyzed the number of correct trials with only 1 response given in an ANOVA, with word category as the repeated-measures factor; early-life adversity and social anxiety disorder as between-subject factors; and age and sex as covariates. We performed post hoc t tests to further investigate interaction effects.
Functional MRI data analysis
We preprocessed and analyzed the data with SPM12 (Wellcome Centre for Human Neuroimaging; www.fil.ion.ucl.ac.uk/spm/). Preprocessing comprised realignment, un-warping using a voxel displacement map derived from the field map, normalization to Montreal Neurological Institute space28 and smoothing with a 6 mm full width at half maximum Gaussian function. At the single-participant level, we defined a general linear model with 1 regressor for each stimulus event and each realignment parameter. A high-pass filter removed frequencies below 1/128 Hz. We estimated a first-order autoregressive model to account for serial correlations, and we calculated a contrast image for social anxiety disorder–related versus neutral stimuli. At the group level, we performed a full factorial ANOVA, with social anxiety disorder and early-life adversity as between-subject factors and age and sex as covariates. We applied a cluster-forming threshold of p < 0.001 (uncorrected) and filtered it for clusters that consisted of 10 or more voxels.
We performed hypothesis-based small-volume correction using a predefined region of interest and assessed significance at a cluster level using a threshold of p < 0.05 (family-wise error [FWE] corrected).29,30 This region of interest included brain regions that were activated in a meta-analysis of attentional control in the context of Stroop tasks (i.e., the inferior frontal junction in the lateral prefrontal cortex, the anterior insula, the anterior midcingulate cortex and the presupplementary motor area).9 We extracted mean contrast estimates from the significant clusters identified in the region of interest analysis and performed posthoc t tests to further investigate interaction effects. To examine linear associations with the severity of early-life adversity, we assessed post hoc Pearson correlation to test for an association between the mean contrast estimates and the total and subscale scores on the Childhood Trauma Questionnaire (separately for participants with and without social anxiety disorder). Results from this correlation analysis underwent Bonferroni correction for multiple testing. We used Fisher z-transformation to test whether correlation coefficients differed between participants with and without social anxiety disorder.
DNA methylation analysis
DNA methylation analysis has been described in more detail in a previous report.31 Briefly, blood samples collected in EDTA tubes (9 mL Monovette; Sarstedt) were available from 84 participants (controls with low early-life adversity: n = 27; controls with high early-life adversity: n = 18; participants with social anxiety disorder and low early-life adversity: n = 20; participants with social anxiety disorder and high early-life adversity: n = 19). DNA was extracted using the QIAamp Blood Maxi Kit (Qiagen) and bisulfite converted using the EZ DNA Methylation Kit (Zymo Research) according to the manufacturers’ instructions. DNA methylation was assessed using the Infinium MethylationEPIC Kit (Illumina).
We performed data processing and analyses in R (version 3.5.1).27 We performed corrections for background and dye bias (Noob),32 probe design bias (BMIQ),33 batch effects (ComBat)34 and cell type composition estimates (Houseman reference data set).35,36
DNA methylation data were filtered down to the 74 CpG sites present on the EPIC array in the NR3C1 gene. For each site, we fitted a linear model, with early-life adversity and social anxiety disorder as main factors (as well as their interaction term) and age and sex as covariates (stats package).27 CpG sites with a p value of less than 0.05 (false discovery rate corrected) for the interaction term were reported as significant and subjected to a conditional process analysis. We performed a moderated mediation analysis to investigate whether NR3C1 DNA methylation (continuous values) might mediate the effects of social anxiety disorder (dichotomous) on brain activity as a function of early-life adversity (dichotomous) using the SPSS implementation of PROCESS (model 8).37 PROCESS is a regression-based analysis tool for mediation, moderation and conditional process analyses. We used the latter application, estimating a conditional indirect effect in a moderated mediation model. We repeated functional MRI analyses in this subcohort as described in the previous section, and used mean contrast estimates from the significant cluster in the moderated mediation analysis as a continuous variable of brain activity.
Results
Sample data
Table 1 shows the sample characteristics for the 4 groups that emerged from the factors early-life adversity and social anxiety disorder, and the effects of social anxiety disorder, early-life adversity and their interaction. We found a significant correlation between total score on the Liebowitz Social Anxiety Scale and total score on the Childhood Trauma Questionnaire (r = 0.35, p < 0.001).
Sample characteristics
Emotional interference
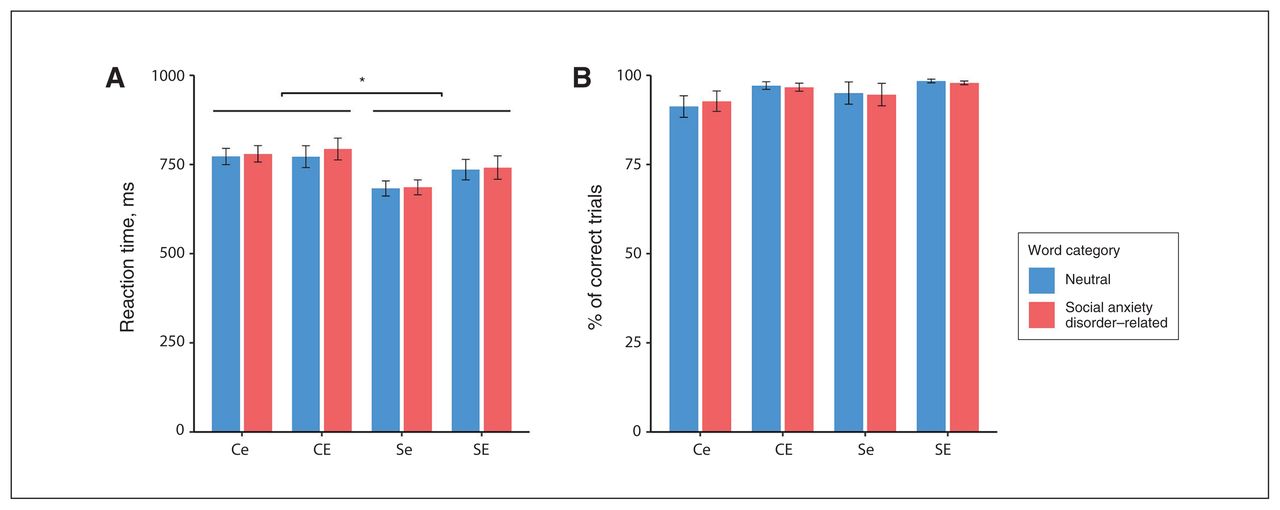
Investigating reaction times for social anxiety disorder–related and neutral words, we found a significant main effect of social anxiety disorder (F1,88 = 6.60, p = 0.012, ηp2 = 0.070) but not of early-life adversity (F1,88 = 0.68, p = 0.41). Further, we found no main effect of word category (F1,88 = 1.30, p = 0.26) and no interaction between word category and social anxiety disorder (F1,88 = 3.88, p = 0.052), word category and early-life adversity (F1,88 = 2.60, p = 0.11) and no significant 3-way interaction (F1,88 = 0.13, p = 0.72). Post hoc t tests showed that participants with social anxiety disorder had significantly shorter reaction times than those without social anxiety disorder (t92 > 2.44, p < 0.017). Figure 1A shows the reaction times for the 2 word categories for each group.

Emotional interference. (A) Mean reaction time (± standard error) for each group. (B) Mean % of correctly answered trials (± standard error) for each group. Ce = control participants with low levels of early-life adversity; CE = control participants with high levels of early-life adversity; Se = participants with social anxiety disorder and low levels of early-life adversity; SE = participants with social anxiety disorder and high levels of early-life adversity. *p < 0.05.
For the number of correct trials, we found no main effect of social anxiety disorder (F1,88 = 0.62, p = 0.43) or early-life adversity (F1,88 = 0.02, p = 0.90). We also found no main effect for word category (F1,88 = 0.75, p = 0.39), and no significant interactions between word category and social anxiety disorder (F1,88.19 = 0.97, p = 0.33), word category and early-life adversity (F1,88 = 2.72, p = 0.10) and no significant 3-way interaction (F1,88 = 1.56, p = 0.22). Figure 1B shows the number of correct trials for the 2 word categories for each group. See Appendix 1, Figure S1, for behavioural data for all 4 word categories.
Functional MRI data
We identified 2 clusters with a significant interaction between social anxiety disorder and early-life adversity in the left and medial prefrontal cortex for the contrast of social anxiety disorder–related and neutral stimuli (Table 2 and Figure 2). The third cluster indicated in Figure 2A (i.e., emerging at an uncorrected threshold of p < 0.001) also partly overlapped with our predefined region of interest but was not significant in our region of interest analysis (pFWE = 0.063). We identified no significant associations with the main effects of social anxiety disorder and early-life adversity. At a descriptive level, we observed no clusters that surpassed the cluster-size threshold of 10 voxels or more outside the a priori region of interest.
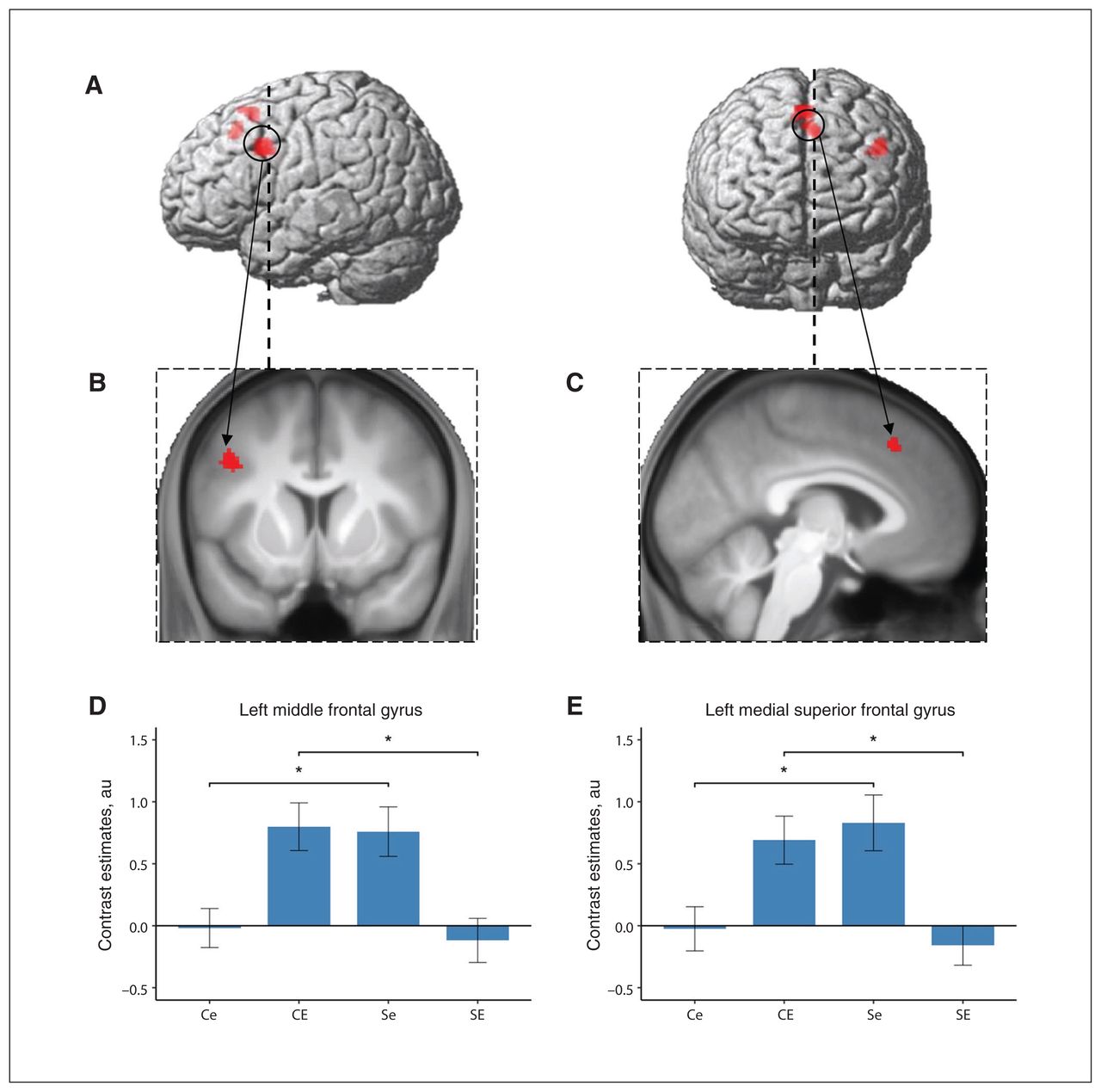
Neural correlates of the contrast “social anxiety–related words versus neutral words” associated with the interaction of social anxiety disorder and early life adversity. Clusters of differential activation at an uncorrected threshold of p < 0.001 and filtered for clusters of 10 voxels or more, rendered on (A) a standard brain and (B) coronal and (C) sagittal slices of the whole sample mean anatomy. (D, E) Mean contrast estimates for each group extracted from the significant clusters identified in the region of interest analysis. Error bars depict standard errors of the mean; *p < 0.05. Ce = control participants with low levels of early-life adversity; CE = control participants with high levels of early-life adversity; Se = participants with social anxiety disorder and low levels of early-life adversity; SE = participants with social anxiety disorder and high levels of early-life adversity.
Neural correlates associated with the interaction of social anxiety disorder and early-life adversity*
The cluster in the left middle frontal gyrus extended to the inferior frontal and precentral gyrus, spanning the border of the ventrolateral and dorsolateral prefrontal cortex and the premotor cortex (Figure 2). Decomposition of the interaction effect in this cluster showed an increased activation difference between social anxiety disorder–related and neutral stimuli in association with social anxiety disorder in participants with low levels of early-life adversity (t53 = −3.06, p = 0.004); in contrast, we found decreased activation differences in association with social anxiety disorder in participants with high levels of early-life adversity (t43 = 3.47, p = 0.001; Figure 2). Exploratory analyses for the other word categories revealed similar trends. The contrasts of “fear-related words versus neutral words” and “negative words versus neutral words” showed no significant effect in participants with low levels of early-life adversity (t53 = −0.75, p = 0.46; t53 = −0.83, p = 0.41), but decreased activation differences in association with social anxiety disorder in participants with high levels of early-life adversity (t43 = 3.45, p = 0.001; t38.7 = 3.54, p = 0.001).
Decomposition of the interaction effect in the medial superior frontal gyrus cluster in the anterior part of the presupplementary motor area extending into the midcingulate cortex showed a similar pattern (Figure 2). For the contrast of “social anxiety disorder–related words versus neutral words,” we found an increased activation difference in association with social anxiety disorder in participants with low levels of early-life adversity (t53 = −2.98, p = 0.004), and a decreased activation difference in association with social anxiety disorder in participants with high levels of early-life adversity (t43 = 3.32, p = 0.002). Likewise, for the contrasts of “fear-related words versus neutral words” and “negative words versus neutral words,” activation differences were increased with respect to social anxiety disorder in participants with low levels of early-life adversity (t53 = −2.51, p = 0.015; t53 = −2.37, p = 0.021) and decreased (at least by trend) in participants with high levels of early-life adversity (t43 = 2.47, p = 0.018; t40= 2.00, p = 0.051; Appendix 1, Figure S2).
Further validation analyses excluding confounds through depressive symptoms, general anxiety or alcohol abuse are reported in Appendix 1.
From a different perspective, Table 3 and Appendix 1, Figure S3, show the correlation of the experienced levels of early-life adversity (total and subcategories) with observed activation differences in the 2 identified clusters. Correlations of emotional neglect with the activity contrasts in both clusters differed significantly between participants with and without social anxiety disorder. Furthermore, correlations of emotional abuse and total score on the Childhood Trauma Questionnaire score with activity contrasts in the left middle frontal gyrus differed significantly between groups.
Pearson correlation of mean contrast estimates extracted from the identified clusters and Childhood Trauma Questionnaire scales
DNA methylation differences and mediation analysis
We investigated DNA methylation at the 74 CpG sites in the NR3C1 gene (Appendix 1, Table S2). DNA methylation at 1 CpG site located in intron 2 of the NR3C1 gene (specifically cg25708981) was associated with the interaction of social anxiety disorder and early-life adversity (pFDR = 0.037). In participants with high levels of early-life adversity, social anxiety disorder was associated with increased methylation levels (t35 = −3.15, p = 0.003; mean ± standard deviation [SD]: controls with high levels of early-life adversity 91.87 ± 1.47%; participants with social anxiety disorder and high levels of early-life adversity 93.34% ± 1.36%). We found no significant difference in DNA methylation associated with social anxiety disorder in participants with low levels of early-life adversity (t45 = 1.97, p = 0.055; controls 92.47 ± 1.51%; social anxiety disorder 91.58% ± 1.57%; Figure 3A). DNA methylation levels at this CpG site were comparable to a previous data set, where the mean (± SD) DNA methylation was 91 ± 6%.38

NR3C1 DNA methylation, neural correlates of the contrast “social anxiety disorder–related words versus neutral words” and their relationship in a moderated mediation model. (A) NR3C1 methylation for each group at site cg25708981. (B) Clusters of differential activation at an uncorrected threshold of p < 0.001 and filtered for clusters of 10 or more voxels rendered on a standard brain. (C) Mean contrast estimates for each group extracted from the significant cluster. (D) Visualization of the direct and indirect effects of social anxiety disorder on brain activity mediated by NR3C1 DNA methylation and moderated by early-life adversity. Error bars depict standard errors of the mean; *p < 0.05. au = arbitrary units; Ce = control participants with low levels of early-life adversity; CE = control participants with high levels of early-life adversity; Se = participants with social anxiety disorder and low levels of early-life adversity; SE = participants with social anxiety disorder and high levels of early-life adversity.
Investigation of brain activity patterns associated with the interaction of early-life adversity and social anxiety disorder in this subcohort (where DNA methylation data were available) identified 1 significant cluster in the left middle frontal gyrus (x, y, z = −40 10 34; F1,78 = 21.05; cluster size 91 voxels, 728 mm3; pFWE (whole brain) = 0.045; Figure 3B). Seventy voxels from this cluster were located in the predefined region of interest (pFWE = 0.005), and 62 of these were within the cluster reported previously for the whole cohort (pFWE < 0.001). Decomposition of this interaction effect revealed an equivalent pattern of activation, as in the cluster for the whole cohort (Figure 3C).
A model of moderated mediation fit the data well (F6,77 = 4.92, p < 0.001; Figure 3D). The analysis demonstrated that NR3C1 DNA methylation (cg25708981) partly mediated the relationship between social anxiety disorder and brain activity in the left middle frontal gyrus as a function of early-life adversity (Table 4).
Mediation of the effect of social anxiety disorder on brain activity by NR3C1 DNA methylation moderated by early-life adversity
Discussion
Investigating the association of social anxiety disorder and early-life adversity with hemodynamic activation in the neural system of attentional control, the key finding of the present study was the identification of 2 brain regions in the left and medial prefrontal cortex that exhibited an interaction between these factors during the processing of disorder-relevant stimuli. Concurrent DNA methylation analysis revealed that social anxiety disorder–related activity differences in the left middle frontal gyrus were partly mediated by NR3C1 DNA methylation as a function of early-life adversity. Importantly, these results provide the first evidence that there might be fundamental differences in the neurobiological processes that underlie social anxiety disorder, depending on the experience of early-life adversity.
Neural interactions of social anxiety disorder and early-life adversity
Although many studies have investigated the neural correlates of social anxiety disorder, findings have been partly contradictory, particularly in prefrontal brain regions.7,8 Although several studies have reported increased activation in the dorsolateral prefrontal cortex in association with social anxiety disorder in response to anxiety-related stimuli,39–41 others have observed decreased activity in the dorsolateral prefrontal cortex.2,42 In the present study, we found that brain activity patterns associated with social anxiety disorder interacted with experiences of early-life adversity. Participants with social anxiety disorder and low levels of early-life adversity showed increased activation to disorder-relevant stimuli compared to healthy controls with equally low levels of early-life adversity; this increased activation did not occur in participants with social anxiety disorder and high levels of early-life adversity. Thus, our findings may suggest contingent differences in early-life adversity as one factor that contributes to the partly contradictory results of previous studies on the neural correlates of social anxiety disorder. Because the prevalence of early-life adversity is associated with economic factors and varies between countries,43 levels of early-life adversity might differ between previous studies depending on their geographic and social settings. Moreover, our findings emphasize the importance of assessing early-life adversity, because it may constitute a relevant moderator of the neural underpinnings of social anxiety disorder. In line with this, previous research has associated early-life adversity with changes in brain activity and suggested early-life adversity as an uncontrolled confound in many neuroimaging studies on psychiatric disorders.14 It is not only that neural alterations might be attributed to social anxiety disorder when they might be more directly related to early-life adversity; it is also that the effects of social anxiety disorder and early-life adversity might interact (as observed in the present study), potentially leading to inconsistent or falsely negative results. Then again, only few studies that investigated brain activity differences in association with early-life adversity have controlled for psychiatric comorbidity.13 To our knowledge, the present study was the first neuroimaging analysis to investigate how neural activations associated with social anxiety disorder might interact with early-life adversity.
Both of the regions we identified coincided with parts of the neural network that is assumed to underlie supervisory attentional control,9 suggesting that the observed alterations were related to this psychological function, which has been causally associated with the maintenance of social anxiety on the basis of empirical data44 and psychological models.45,46 In contrast, no brain regions outside of the attentional control network exhibited any interaction effect. Our data suggest that participants with social anxiety disorder and low levels of early-life adversity and controls with high levels of early-life adversity engaged these brain regions more during attentional control in the context of negative emotional processing, and most prominently in the context of social anxiety disorder–related information processing. This finding might be indicative of a compensatory mechanism, with increased attentional effort as a consequence of increased automatic attention capture through this type of emotional information. However, we did not observe such increased neural responsiveness in participants with social anxiety disorder and high levels of early-life adversity, whose activity patterns resembled control participants with low levels of early-life adversity in the identified regions. This initially counterintuitive reversal of increased activity in participants with social anxiety disorder and early-life adversity was not specific to social anxiety disorder–related stimuli; it was also present for fear-related and generally negative stimuli. This finding might indicate a more general alteration in neural responsivity to negative emotional information compared to neutral information in participants with social anxiety disorder and high levels of early-life adversity.
A recent study on the interrelations of social anxiety, dissociative symptoms and early-life adversity might serve as a starting point for the interpretation of this unexpected finding. It demonstrated that early-life adversity moderated the association between social anxiety and dissociative symptoms:47 a positive correlation was observed in participants with high levels of early-life adversity, but not in participants with low levels of early-life adversity. Dissociation, which describes the feeling of emotional or physical detachment, is regarded as a defence mechanism to avoid unpleasant emotional states, typically occurring after the experience of childhood trauma.48 From the perspective of this line of research, the observed pattern with a generally reduced neural responsiveness to negative emotional stimuli relative to neutral stimuli might be related to a higher degree of dissociation in participants with social anxiety disorder and high levels of early-life adversity.
Recent research on the neural correlates of early-life adversity revealed effects that were selective for different subtypes of early-life adversity: emotional but not physical maltreatment has been associated with increased activity in the ventrolateral and dorsolateral prefrontal cortex.49 Concordantly, we observed significant positive correlations between brain activity and early-life adversity in participants without social anxiety disorder, for the emotional aspects of early-life adversity, but not the physical aspects. Interestingly, the negative correlation between brain activity and early-life adversity in participants with social anxiety disorder was also correlated with an emotional facet of early-life adversity (i.e., emotional neglect), but none of the physical subtypes of early-life adversity. This finding was also in line with previous research, indicating a more prominent contribution of emotional trauma to the psychopathology of social anxiety disorder compared to physical or sexual maltreatment.50 Therefore, the interaction observed in the present study might be more strongly related to emotional rather than physical or sexual aspects of childhood trauma.
In contrast to the interaction between social anxiety disorder and early-life adversity at the neural level, participants’ response behaviour in the emotional Stroop task evidenced a main effect of social anxiety disorder driven by faster reaction times in participants with social anxiety disorder. It is conceivable that this behavioural effect was based on increased motivation to perform well in the experimental task, elicited by a central feature of social anxiety disorder pathology — namely feared scrutiny by others, such as the experimenters outside the scanner room.
Epigenetic mediation of the interaction between social anxiety disorder and early-life adversity
It has been suggested that epigenetic modifications of the gene encoding the glucocorticoid receptor, NR3C1, physiologically mediate the long-term effects of early-life adversity on the functioning of the hypothalamic–pituitary–adrenal axis.51 We observed that DNA methylation at 1 CpG site in NR3C1 mediated differential activity in the left middle frontal gyrus associated with social anxiety disorder as a function of early-life adversity. Participants with social anxiety disorder and low levels of early-life adversity showed decreased NR3C1 DNA methylation compared to control participants with equally low levels of early-life adversity. In contrast, in participants with high levels of early-life adversity, NR3C1 DNA methylation patterns were reversed. If one cautiously assumed that lower DNA methylation levels were associated with increased gene expression of NR3C1, the corresponding increase in glucocorticoid receptors might be correlated with increased sensitivity to glucocorticoids. Binding of released glucocorticoids to the glucocorticoid receptor allows for regulation of the expression of glucocorticoid-responsive genes52 and plays a critical role in the negative feedback of the hypothalamic–pituitary–adrenal axis.53 The CpG site reported here — cg25708981, within intron 2 of the NR3C1 gene — has previously been found to be hypomethylated in buccal cells in response to exposure to a synthetic glucocorticoid.54 However, previous studies have shown a reduced activity in the hypothalamic–pituitary–adrenal axis in response to social stress in individuals with a history of early-life adversity.55 In this regard, increased NR3C1 DNA methylation in those with social anxiety disorder and a history of early-life adversity might represent an adaptation to repeated and enduring exposure to increased cortisol levels and be part of the underlying molecular basis for the reduced reactivity of the left middle frontal gyrus to negative emotional stimuli.
Limitations
The novel findings of our work provide a foundation for several strains of research to validate and extend our findings.
The absence of social anxiety disorder–associated behavioural correlates of altered attentional control in the present study somewhat limited the functional interpretation of our findings. The identified brain regions fit well with our a priori hypothesis of activity differences in the attentional control system, although we cannot rule out the possibility that other regions outside this network were involved. In future studies, designs with greater power to elicit behavioural correlates of altered attentional control in those with social anxiety disorder and early-life adversity (e.g., using multimodal stimuli more closely approximating real-life social threat56–58) are needed to validate our interpretations and clarify the further functional implications of the observed effects. Beyond behavioural correlates, longitudinal studies that include interventions to modulate attentional alterations (e.g., attention bias modification training59 or noninvasive brain stimulation60) may help to explore the functional and clinical relevance of our results. Moreover, in the present study the variance for physical and sexual adversity was low, limiting the generalizability of our results to different types of early-life adversity.
The potential link of diminished emotional reactivity and dissociative symptoms in those with social anxiety disorder and high levels of early-life adversity represents another starting point for further research. This link appears to be very relevant, because symptoms of dissociation are present in many psychiatric disorders and have been connected to higher burden of illness and poorer treatment response.61
At the epigenetic level, we measured DNA methylation in whole blood and not in brain tissue, where it can be assumed to be more directly linked to brain activity. Nevertheless, the association of early-life adversity with differential NR3C1 DNA methylation has been demonstrated not only in peripheral blood51 but also in buccal cells62 and the hippocampus.63 Furthermore, if changes in NR3C1 DNA methylation are provoked by aberrant cortisol levels,54 the effects would not be limited to blood cells. Because cortisol passes the blood–brain barrier, it can be expected to have a similar effect on brain tissue, where it would be more directly related to the experience of stress and anxiety. Nevertheless, the observed differences in NR3C1 DNA methylation were relatively modest and the functional relevance of the identified CpG site needs to be clarified in studies with gene expression analyses. In addition, inclusion of further explanatory variables such as genotype data and subsequent trauma load might further improve the interpretability of the present findings. Moreover, because stress exposure during different time windows in childhood and adolescence may lead to different outcomes,64 future studies should assess the exact timing of early-life adversity to test whether it alters the moderating effect of early-life adversity on NR3C1 DNA methylation and the brain activity patterns associated with social anxiety disorder. Similarly, the question arises whether adverse events later in life might also interact with the observed effects.
Investigating different aspects of the neurobiological mechanisms that underlie social anxiety disorder, we performed various statistical analyses in this study. Although we ensured multiple testing correction within separate analyses, such a comprehensive approach is generally accompanied with an increased risk of type I errors. The key findings of the present study need to be replicated in independent cohorts.
Conclusion
The present study provides the first evidence for essential differences in the neurobiological processes associated with social anxiety disorder, depending on a history of early-life adversity. Our data suggest that long-lasting effects of early-life adversity might be encoded in NR3C1 DNA methylation patterns and manifest in altered brain activity in the attentional control network during confrontation with anxiety-related stimuli.
Supplementary Information
Acknowledgements
The authors thank Danuta Altpaß, Gisbert Farger and Katia E. Ramadori for technical assistance. They also thank Cieslik and colleagues for sharing the results of their meta-analysis. They acknowledge support from Deutsche Forschungsgemeinschaft (DFG, NI 1332/6-1, FA 361/25-1, KR 4398/5-1) and the Open Access Publishing Fund of the University of Tübingen. The authors also wish to express their appreciation to all participants.
Footnotes
Competing interests: None declared.
Contributors: A. Fallgatter, V. Nieratschker and B. Kreifelts conceived and designed the study. A. Wiegand, M. Munk and S. Drohm recruited the participants and contributed to data acquisition. J. MacIsaac and M. Kobor contributed to the generation of the DNA methylation data. A. Wiegand and B. Kreifelts analyzed the data. A. Wiegand wrote the manuscript. M. Munk, S. Drohm, A. Falgatter, J. MacIsaac, M. Kobor, V. Nieraschker and B. Kreifelts revised the manuscript substantially. All authors approved the final version to be published and can certify that no other individuals not listed as authors have made substantial contributions to the paper.
- Received April 23, 2021.
- Revision received August 10, 2021.
- Accepted October 10, 2021.
This is an Open Access article distributed in accordance with the terms of the Creative Commons Attribution (CC BY-NC-ND 4.0) licence, which permits use, distribution and reproduction in any medium, provided that the original publication is properly cited, the use is noncommercial (i.e., research or educational use), and no modifications or adaptations are made. See: https://creativecommons.org/licenses/by-nc-nd/4.0/
References
In this issue

{kind=link}
{kind=link}
{kind=link}
Article tools



Related Articles
Cited By...
- No citing articles found.