

Article Figures & Tables
Figures
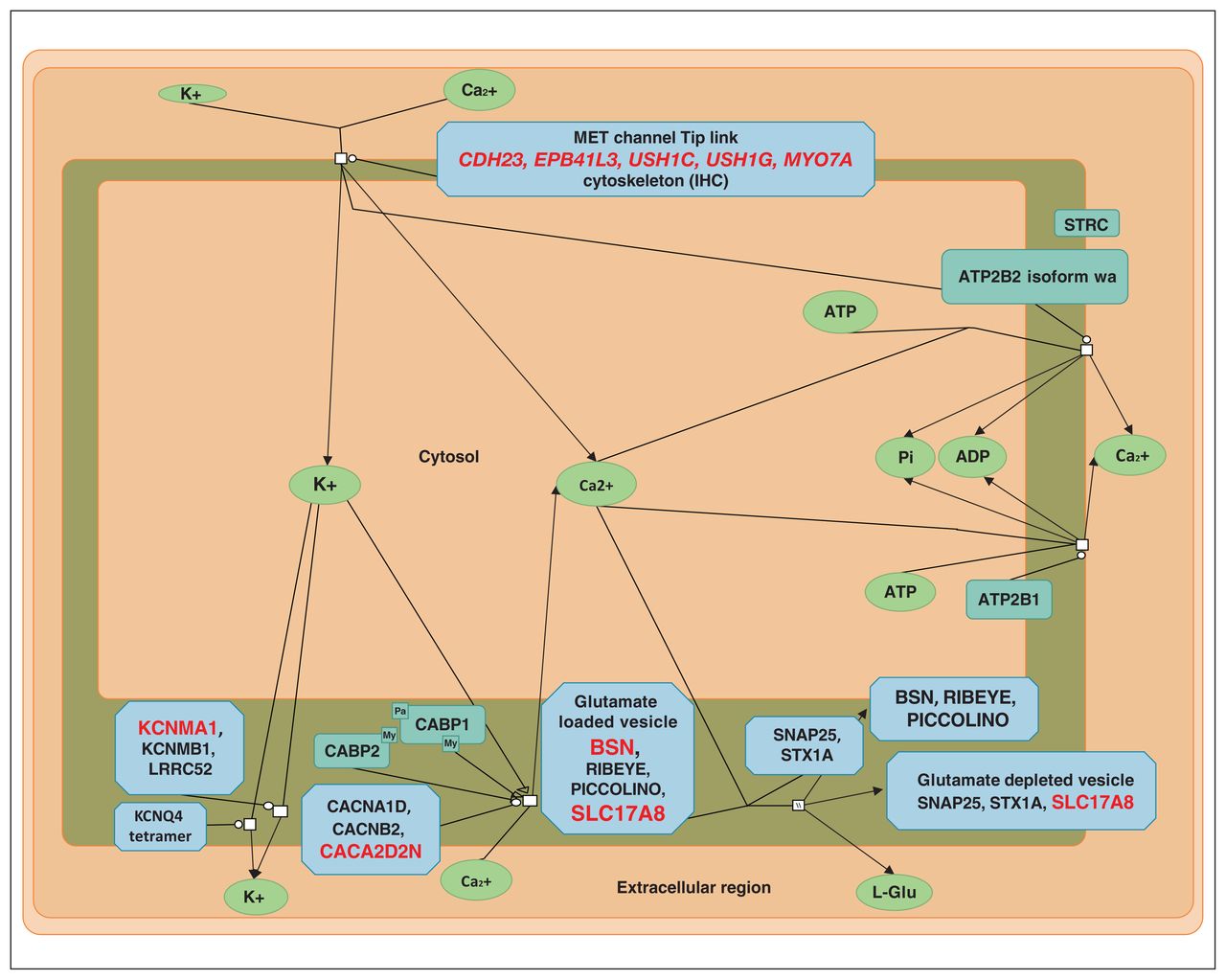
- Figure 1
Sensory processing of sound by inner hair cells of the cochlea (modified from reactome.org). Proteins coded by genes with variants found in the present study are shown in red.
Tables
Family code Members, n [M/F] (n = 124) Tourette syndrome, n (n = 40) Other tic disorders, n (n = 40) Cosegregation probability P(seg) Healthy controls, n (n = 44) Comorbidities A 7 [4/3] 3 1 0.0625 3 ANX B 8 [5/3] 3 2 0.0156 3 ADHD, ANX, MDD, OCD C 9 [4/5] 2 4 0.0078 3 ANX D 7 [5/2] 2 2 0.0156 3 ANX, MDD, OCD E 7 [4/3] 2 2 0.0625 3 – F 11 [7/4] 4 4 0.0078 3 – G 4 [2/2] 1 2 0.25 1 ANX H 6 [4/2] 2 3 0.125 or 0.125* 1 ADHD, ANX, OCD I 6 [4/2] 2 2 0.0625 2 ANX, MDD, OCD J 5 [2/3] 1 2 0.25 2 ANX, MDD, OCD R 6 [4/2] 1 3 0.125 2 ANX S 6 [2/4] 4 0 0.125 2 MDD, OCD T 9 [4/5] 2 3 0.125 or 0.25* 4 OCD U 6 [5/1] 2 2 0.0625 2 – W 14 [7/7] 4 4 0.000976 6 – X 5 [2/3] 2 1 0.125 2 ADHD Y 8 [4/4] 3 3 0.03125 2 MDD, OCD ADHD = attention-deficit and hyperactivity disorder; ANX = anxiety disorder; F = female; M = male; MDD = major depression disorder; OCD = obsessive– compulsive disorder.
↵* In families H and T, 2 segregation patterns were analyzed (see Appendix 1, available at www.jpn.ca/lookup/doi/10.1503/jpn.220206/tab-related-content).
Location/type of variant Number of variants Ultra-rare Rare Uncommon Intronic 206 288 776 Exonic/missense 44 24 54 Noncoding exon 6 5 25 5′ UTR 8 4 9 3′ UTR 11 7 20 Intergenic 5 6 15 Splicing 4 2 5 Exonic/synonymous 3 1 2 Upstream 2 0 2 Downstream 0 0 1 Nonsense 0 0 1 MAF = minor allele frequency; UTR = untranslated region.
↵* Included are variants found in genes called in at least 2 families (ultra-rare variants, MAF < 0.1%), at least 3 families (rare variants, MAF < 1%), or at least 5 families (uncommon variants, MAF < 5%).
- Table 3
Rare and ultra-rare variants with the CADD score > 30 segregating with the disease in any single family
Position and change Gene Type of variant CADD MAF, % ACMG classification (ACMG criteria met) Family, P(obs)* chr7:106112481 G/A SYPL1 Stop gain 41.0 0 Likely pathogenic (PP3, PM2) Y, 0.00278 chr9:38396880 C/T ALDH1B1† Stop gain 40.0 0.203 VUS with pathogenic evidence (PM2, PP3) T, 0.02225 chr18:12702512 G/A CEP76 (PSMG2) Stop gain 40.0 0 VUS with pathogenic evidence (PM2, PP3) X, 0.01113 chr12:2855156 C/T TEX52 (ITFG2) Stop gain 37.0 0.166 VUS with pathogenic evidence (PM2, PP3) H, 0.01113 chr11:113697354 C/T TMPRSS5 Stop gain 36.0 0.031 Likely pathogenic (PVS1, PM2) U, 0.00556 chr1:117120939 G/T TRIM45 Stop gain 36.0 0.182 Benign (BS1, BS2, PVS1) J, 0.02225 chr3:149073278 C/A HLTF Stop gain 36.0 0.001 VUS with pathogenic evidence (PM2, PP3) I, 0.002781 chr4:81459515 C/T RASGEF1B Splicing 35.0 0.030 Pathogenic (PVS1, PM2) J, 0.02225 chr14:96264526 C/T BDKRB1 Stop gain 35.0 0.250 Benign (BS1, BS2, BP4) I, 0.002781 chr3:53855341 G/A IL17RB Missense 35.0 0.726 Benign (BS1, BS2, BP6) A, 0.00556 chr7:2569000 G/A IQCE Splicing 34.0 0.024 Likely pathogenic (PVS1, PM2) S, 0.01113 chr10:94687805 T/A CYP2C18 Stop gain 34.0 0.316 Benign (BA1, BP4) J, 0.02225 chr4:88417579 G/T HERC6 Missense 34.0 0.396 Benign (BS1, BS2, BP1, PP3) J, 0.02225 chr17:69026976 C/T ABCA9 Missense 34.0 0 VUS with pathogenic evidence (PM2, PP3) U, 0.00556 chr16:2317763 T/A ABCA3 Missense 33.0 0.501 Benign (BS1, BS2, PP3, PP5) T, 0.02225 chr3:132601106 C/T ACKR4 (ACAD11, NPHP3-ACAD11) Stop gain 33.0 0.003 Uncertain significance (PM2, BP4) I, 0.002781 chr12:100396328 A/C SLC17A8‡ Splicing 33.0 0 Pathogenic (PVS1, PM2) T, 0.02225 chr12:101319592 G/A UTP20 Missense 32.0 0 Uncertain significance (PM2, PP3, BP1) G, 0.01113 chr12:1885984 G/A CACNA2D4 Missense 32.0 0 VUS with pathogenic evidence (PM2, PP3) H, 0.01113 chr3:49004877 C/T P4HTM Missense 32.0 0.013 Uncertain significance (PM2, BP4) T, 0.02225 chr17:10631666 T/C MYH3 Missense 32.0 0.025 VUS with pathogenic evidence (PM2, PP3) J, 0.02225 chr15:101324994 G/A PCSK6 Missense 32.0 0.059 Uncertain significance (PM2, BP1) J, 0.02225 chr16:4883689 G/A PPL Missense 32.0 0.156 Uncertain significance (PM2, PP3, BP1) J, 0.02225 chr17 28385146 T/C SARM1 Missense 32.0 0.291 VUS with pathogenic evidence (PM2, PP3) G, 0.01113 chr16:89727323 C/T ZNF276 Missense 32.0 0.969 Benign (BS1, BS2, BP6) R, 0.01113 chr12:101677317 G/A MYBPC1 Missense 32.0 0.001 VUS with pathogenic evidence (PM2, PP3) U, 0.00556 chr12:111803962 G/A ALDH2† Missense 32.0 0.003 Benign (BA1, BP6, BS4, PP3) U, 0.00556 chr12:94279637 G/A PLXNC1 (CEP83) Missense 32.0 0 Uncertain significance (PM2, PP3, BP1) G, 0.01113 chr12:57244143 G/A STAC3 Missense 31.0 0.024 Uncertain significance (PM2) G, 0.01113 chr6:39861001 A/G DAAM2 Missense 31.0 0.028 Likely pathogenic (PM2, PM1, PP3) H, 0.01113 chr8:99999384 G/A RGS22 Missense 31.0 0.046 Benign (BA1, BP6, PP3) S, 0.01113 chr21:46114063 G/A COL6A2 Missense 31.0 0.090 VUS with pathogenic evidence (PM1, PM2) H, 0.01113 chr2:108499560 A/G GCC2 Missense 31.0 0.876 Benign (BA1, BP1, PM2, PP3) H, 0.01113 chr3:49652822 G/C BSN‡ Missense 31.0 0 VUS with pathogenic evidence (PM2, PP3) D, 0.00139 chr16:80549649 C/G DYNLRB2 (AC105411.1AC108097.1) Missense 31.0 0 VUS with pathogenic evidence (PM2, PP3) I, 0.002781 chr7:107916945 T/C DLD† Missense 30.0 0 VUS with pathogenic evidence (PM2, PP3) J, 0.02225 chr9:37541696 G/A FBXO10 (AL513165.2) Missense 30.0 0 VUS with pathogenic evidence (PM2, PP3) T, 0.02225 ACMG = American College of Medical Genetics and Genomics; CADD = combined annotation dependent depletion; MAF = minor allele frequency; VUS = variant of unknown significance.
↵* P(obs) = probability of cosegregation of a rare variant with a CADD > 30 in a given family, Bonferroni corrected to 17 families: P(obs) = P(seg) × (P(var) × 17. P(var) = 1 – Πn1 (1 – MAFn) n = 67 – overall number of variants with CADD > 30.
↵† Genes encoding dehydrogenases influencing oxidoreductase activity.
↵‡ Genes involved in sensory processing of sound by hair cells of the cochlea.
Position change Gene* Location/ type of variant MAF, % CADD ACMG classification (ACMG criteria met) Fam, P(obs)† 19:53803962 G/A NLRP12 Exonic/missense 0.01 23.3 Likely benign (BS2) G, I, 0.000211 5:76482747 A/C IQGAP2 Intronic 0.08 15.9 Benign (BS1, BS2) E, X, 0.000842 4:109086168 T/A COL25A1 Intronic 0.10 19.0 Likely benign (BS1) H, J, 0.004208 1:223300619 G/A SUSD4 Intronic 0.20 16.4 VUS with benign evidence (BP4) J, X, 0.008415 12:120368947 C/A MSI1 Intronic 0.33 16.4 Benign (BS1, BS2, BP4) J, R, 0.013885 1:31727178 G/A ADGRB2 3′ UTR 0.60 13.2 Benign (BS1, BS2) B, G, 0.003151 12:10723010 G/T YBX3 Exonic/missense 0.72 22.2 Likely benign (BS1, BP4) J, T, 0.030294 11:111020375 C/A AP003973.4 Intergenic 0.73 14.0 Benign (BS1, BS2, BP4, BP7) E, S, 0.007679 6:101375096 A/G GRIK2 Intronic 0.73 20.5 Benign (BS1, BS2) D, G, 0.003833 12:97272797 T/A LINC02409 Intronic/splice 0.75 14.7 Benign (BA1, BP4, BP7) G, X, 0.031556 7:713068 C/T PRKAR1B 5′ UTR 0.75 10.5 Benign (BS1, BS2, BP4) A, J, 0.015778 1:121379326 G/A SRGAP2C Intronic 0.91 15.5 Benign (BA1, BP4) J, X, 0.038288 10:74875378 T/C KAT6B Intronic 0.95 11.0 Benign (BS1, BS2, BP4) J, X, 0.039972 6:17649263 G/A NUP153 Exonic/missense 0.99 27.7 Benign (BS1, BS2, BP1, PP3) C, J, 0.002599 ACMG = American College of Medical Genetics and Genomics; CADD = combined annotation dependent depletion; MAF = minor allele frequency; UTR = untranslated region; VUS = variant of unknown significance.
↵* Several genes had more than 1 ultra-rare variant with putatively severe consequences. Seven genes (BSN, FLG-AS1, FN1, NLRP12, PTPN14, RBL2, and SNX19) had 2 missense variants each, with a third variant in BSN located in 3′ UTR. Three out of 4 variants located in TTN were missense. Eight genes had 1 missense and 1 3′ UTR or 5′ UTR variant: AC046130.1, GAPVD1, IGSF3, NOL4L, ROBO2, SEMA4A, TMEM63B, TPRG1.
↵† P(obs) – probability of cosegregation of a particular variant in a given 2 families, Bonferroni corrected to 136 possible pairs of families: P(obs) = P(seg_fam1) × P(anyvar) × P(seg_fam2) × P(var) × 136. P(any var) = 0.99 – probability of a variant in family 1 with CADD > 10 and MAF < 0.01. P(var) = MAF – the probability of observing the particular variant in the second family P(var).
- Table 5
Top significantly enriched categories (LogP value < −4) with variants associated with Tourette syndrome
Term ID Description LogP Genes in which variants were found MAF below, %* GO:0098609 Cell-cell adhesion −13.56 CDH23†‡, NLGN1†, PTPRT†, ROBO2†‡, ADGRL3‡, CD44, CLSTN2, CNTN4‡, CTNNA3, FN1‡, GNAS, LPP, NRXN3, PKHD1‡, PKP4, PTPRD, SDK1‡, TENM2‡ 5.0 GO:0000902 Cell morphogenesis −7.93 CDH23†‡, EPHB1†, NLGN1†, ROBO2†‡, SEMA3A†‡, AUTS2, CNTN4‡, FN1‡, NCAM1‡, NRG1, NRXN3, PKHD1‡, PLXNC1‡, USH2A‡ 5.0 GO:0034329 Cell junction assembly −7.75 ERBB4, BSN‡, PKP4†, PTPRD†, ADGRL3‡, NRXN1, CDH22, EPB41L3‡, FN1‡, CLDN14 0.1 GO:0034330 Cell junction organization −7.70 EPHB1†, NLGN1†, ADGRL3‡, DLC1‡, ERBB4, ERC2, FN1‡, NRG1, PKHD1‡, PKP4, PTPRD, SDK1‡ 5.0 GO:0098609 Cell-cell adhesion −7.57 TENM2, CDH22, CDH23‡, CNTN4‡, ROBO2†‡, PTPRD†, ADGRL3‡, CLDN14, PIK3CG, NRXN1, NRXN3, PKP4†, FN1‡ 0.1 R-HSA-9662360 Sensory processing of sound by inner hair cells of the cochlea −6.94 KCNMA1, EPB41L3‡, CDH23‡, CACNA2D2‡, BSN‡, SLC17A8‡ 0.1 GO:0007420 Brain development −6.77 SATB2, ERBB4, MEIS2, RARB, ROBO2†‡, ALK‡, PRKG1†, CDH22, NDRG2‡, ADGRL3‡, EPHB1, ZNF148, SLC17A8‡, CNTN4‡ 0.1 GO:0016358 Dendrite −6.56 CACNA1C†, EPHB1†, NLGN1†, SEMA3A†‡, ANKS1B, CLSTN2, GIGYF2‡, KCNIP4, MAGI2, SLC4A10, SLC8A1, TENM2‡ 5.0 GO:0007420 Brain development −6.50 EPHB1†, ROBO2†‡, SEMA3A†‡, MACROD2†, ADGRL2, ADGRL3‡, CNTN4‡, DLC1‡, ERBB4, NRG1, PLCB1, RARB, SLC4A10, SLC8A1 5.0 GO:0008038 Neuron recognition −6.44 OPCML†, ROBO2†‡, NTM†, CNTN4‡ 5.0 GO:0030424 Axon −6.41 EPHB†, ROBO2†‡, SEMA3A†‡, ADGRL3‡, AUTS2, CNTN4‡, ERC2, NRG1, SLC4A10, SLC8A1, TENM2‡, USH2A‡ 5.0 GO:0000902 Cell morphogenesis −6.06 CDH22, CDH23†‡, EPHB1, KLF7, NLGN1, NRXN3, NTN4, PLXNC2‡, ROBO2†‡, SEMA3A†‡ 1.0 GO:0098609 Cell-cell adhesion −5.90 CDH22, CDH23†, GNAS†, NLGN1, NRXN3, PKP4, PTPRD, ROBO2†‡, TENM2†‡ 1.0 GO:0051963 Regulation of synapse assembly −5.90 NRXN1, EPHB1, SEMA4A‡, ROBO2†‡, PTPRD†, COLQ 0.1 GO:0007626 Locomotory behaviour −5.85 BTBD9†, CDH23†‡, FGF12, GIGYF2‡, NAV2‡, NRG1, SLC4A10 5.0 GO:0005509 Calcium ion binding −5.85 CDH23†‡, EYS†, ADGRL3‡, CLSTN2, FSTL5, KCNIP4, LTBP1‡, PLCB1, SLC8A1, STAB2‡, TENM2‡, TLL2‡ 5.0 GO:0040007 Growth −5.75 EYS†, SEMA3A†‡, AUTS2, ERBB4, GIGYF2‡, GNAS, MAGI2, RARB, SLC4A10 5.0 GO:0007167 Enzyme-linked receptor protein signalling pathway −5.72 EPHB1†, PTPRT†, ANKS1B, ERBB4, FGF12, GIGYF2‡, LTBP1‡, MAGI2, NRG1, PLCB1, PTPRD 5.0 GO:0007420 Brain development −5.65 ADGRL2, ALK‡, CDH22, EPHB1, ERBB4, MACROD2†, PRKG1, RARB†, ROBO2†‡, SEMA3A†‡, SRGAP2C 1.0 GO:0051962 Positive regulation of nervous system development −5.47 FN1‡, SEMA4A‡, MAP3K13, ROBO2†‡, PLXNC1‡, PTPRD†, EPHB1, NRXN1 0.1 GO:0120035 Regulation of plasma membrane bounded cell projection organization −5.36 ALK‡, NLGN1, PLCE1‡, PLXNC1‡, PTPRD, ROBO2†‡, SEMA3A†‡, SRGAP2C, TENM2†‡ 1.0 GO:0098858 Actin-based cell projection −5.34 CDH23†‡, EPHB1†, NLGN1†, CD44, IQGAP2‡, TENM2‡, USH2A‡ 5.0 GO:0034330 Cell junction organization −5.27 CDH22, EPHB1, ERBB4, ERC2, NLGN1, PKP4, PTPRD, SRGAP2C 1.0 GO:0050804 Modulation of chemical synaptic transmission −5.21 BTBD9†, EPHB1†, NLGN1†, CLSTN2, CNTN4‡, ERC2, PLCB1, PTPRD, SLC4A10 5.0 GO:0016324 Apical plasma membrane −4.89 KCNMA1†‡, CD44, FN1‡, GNAS, PARD3B, PKHD1‡, SLC4A10, USH2A‡ 5.0 GO:0005539 Glycosaminoglycan binding −4.89 COL25A1†, CD44, FN1‡, HK1‡, NAV2‡, STAB2‡ 5.0 GO:0050885 Neuromuscular process controlling balance −4.67 NRXN1, RBFOX1, CAMTA1, CDH23‡ 0.1 GO:0016358 Dendrite development −4.16 EPHB1, KLF7, PRKG1, SEMA3A†‡ 1.0 GO:0045932 Negative regulation of muscle contraction −4.11 PRKG1†, KCNMA1, PIK3CDG 0.1 WP2118 Arrhythmogenic right ventricular cardiomyopathy −4.10 CACNA1C†, CACNA2D3†, CTNNA3, SLC8A1 5.0 GO:0098858 Actin-based cell projection −4.07 CDH23†, EPHB1, NLGN1, TENM†‡, WWOX 1.0 GO:0043408 Regulation of MAPK cascade −4.01 EPHB1, ERBB4, ALK‡, MAP3K13, PIK3CG, FN1‡, NDRG2‡, NLRP12‡, PLCE1‡, MAPKBP1‡ 0.1 MAF = minor allele frequency.
↵* Analyses were performed separately on gene sets with ultra-rare variants (MAF below 0.1%) occurring in at least 2 families, rare variants (MAF below 1%) occurring in at least 3 families, and uncommon variants (MAF below 5%) occurring in at least 4 families.
↵† Genes with variants found in all patients in at least n + 1 families, where n is the threshold value selected for a given MAF (i.e., 2 for 0.1%, 3 for 1% and 4 for 5%) threshold.
↵‡ Genes with missense and/or splicing variants.
- Table 6
Genes identified as being associated with Tourette syndrome that were previously implicated in psychiatric and neurologic disorders
Disorder/phenotype Genes Autism spectrum disorder NRXN3, NLGN1, PIK3CG, EPB41L3, CACNA1C Schizophrenia ERBB4, NRXN1 Attention-deficit/hyperactivity disorder ADGRL3, PTPRD Obsessive–compulsive disorder PTPRD Restless legs syndrome PTPRD, BTBD9 Deafness CLDN14, CDH23, Speech/sound disorders ROBO2, KCNMA1 Alzheimer disease CTNNA3 Amyotrophic lateral sclerosis ERBB4 Developmental delay KCNMA1, CELF2 Other neurological COL25A1, ZBTB20, CELF2, CACNA1C - Table 7
Variants in genes involved in the sensory processing of sound by inner hair cells of the cochlea*
Gene Family D Family E Family G Family H Family I Family J† Family R Family T BSN ur, 31, m – – – – ur,15, UTR
ur, 11, m– – CACNA2D2 – – – – – ur, 19, i
ur, 15, i– ur, 22, m CDH23 ur, 22, i
r, 17, i
r, 17, i
ur, 14, ir, 20, i
uc, 18, i
uc, 16, i
r, 13, i– uc, 21, i
uc, 18, ir, 23, m ur, 21, i
ur, 20, m
r, 16, i
ur, 15, i
r, 14, i
r, 11, i
r, 10, i– – EPB41L3 – – – – – ur, 13, s ur, 24, m – KCNMA1 – uc, 12, i
uc, 11, iur, 12, i r, 12, i
uc, 13, iuc, 17, i
uc, 16, i
ur, 14, iuc, 17, i
uc, 13, s– – SLC17A8 – – ur, 10, i – – – – ur, 33, sp CADD = combined annotation dependent depletion; i = intronic; m = missense; MAF = minor allele frequency; r = rare (MAF 0.1%–1%); s = synonymous; sp = splicing; uc = uncommon (MAF 1%–5%); ur = ultra-rare (MAF < 0.1%); UTR = untranslated region.
↵* Numbers indicate CADD score.
↵† The high number of variants found in Family J was partially due to the close kinship among all 3 affected family members (father and 2 siblings).
In this issue

{kind=link}
Article tools



Related Articles
Cited By...
- No citing articles found.